

Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...
Топ:
Основы обеспечения единства измерений: Обеспечение единства измерений - деятельность метрологических служб, направленная на достижение...
Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Интересное:
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
КОКОРИНА Н. В.
УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ
«ИЗБРАННЫЕ ГЛАВЫ КУРСА «БИОЛОГИЯ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ»
для студентов второго курса специальности «медицинская экология»
Минск, 2006

Автор:
Н. В. Кокорина, доцент, к.м.н.
Рекомендовано Советом факультета экологической медицины МГЭУ°им.°А.Д.°Сахарова в качестве инструктивно-методических материалов (протокол № ___ заседания Совета факультета экологической медицины от _________).
Содержание:
Лекция № 1. Молекулярно-генетические механизмы
Индивидуального развития 4
Лекция № 2. Гаметогенез и оплодотворение 10
Лекция № 3. Гаструляция 18
Лекция № 4. Развитие различных классов позвоночных 28
Лекция № 5. Развитие человека 42
Приложение 1 54
Лекция № 3. Гаструляция
Темпы и ритм дробления регулируются многими факторами и зависят от вида животного. Характер дробления определяется количеством желтка и разным распределением его в яйцеклетке. Желток мешает закономерным движениям составных частей ядра и цитоплазмы, и выстраивания веретена деления. Существует зависимость между количеством и распределением желтка и положением борозд дробления (правила О. Гетрвига):
1) клеточное ядро стремится расположиться в центре свободной от желтка цитоплазме;
2) веретено клеточного деления стремится расположиться по направлению свободного от желтка пространства цитоплазмы, поэтому борозда каждого последующего дробления перпендикулярна борозде предыдущего;
3) плоскость дробления всегда перпендикулярна оси веретена деления, поскольку компоненты веретена деления воздействуют на кортикальный слой и стимулируют появление борозды дробления в определенной плоскости.
|
|
У маложелтковых (олиголецитальных) яйцеклеток дробление полное (голобластическое), равномерное. Яйцеклетки с умеренным содержанием желтка (мезолецитальные) дробятся полно, но неравномерно, так как желток, сосредоточенный на вегетативном полюсе, замедляет темпы дробления.
Яйцеклетки с очень высоким содержанием желтка (полилецитальные) дробятся частично и неравномерно (меробластический тип дробления). У плацентарных млекопитающих и человека яйцеклетка содержит небольшое количество желтка, распределенного в цитоплазме равномерно. Такая яйцеклетка называется вторичной олигоизолецитальной. Дробление голобластическое, неравномерное и асинхронное, что связано с особенностями строения бластомеров. В зависимости от формы и перемещения бластомеров различают:
- голобластическое радиальное дробление (ланцетник) – бластомеры правильной сферической формы, расположены один над другим;
- голобластическое спиральное (черви, моллюски) – бластомеры отличаются по форме и размерам, в ходе дробления поворачиваются вокруг оси веретена деления;
- голобластическое билатеральное (асцидии) – дробление осуществляется вокруг единственной плоскости симметрии, и половины зародыша являются зеркальным отражением друг друга;
- голобластическое анархическое – бластомеры располагаются неправильными цепочками (голотурии);
- голобластическое чередующее (млекопитающие) – первое деление меридиальное, при втором один бластомер делится меридиально, второй – экваториально. При этом темпы деления клеток неодинаковы, поэтому на определенных этапах дробления зародыш содержит нечетное количество бластомеров.
Для процесса дробления характерна компактизация. Сначала в бластуле млекопитающих бластомеры располагаются рыхло, затем они сближаются и образуют компактный шар, поскольку между клетками устанавливаются плотные контакты. Клетки, лежащие на поверхности бластоцисты, образуют трофобласт, а сгруппированные внутри – эмбриобласт. Между бластомерами, изолированными внутри бластоцисты устанавливаются щелевые контакты, которые позволяют малым молекулам и ионам перемещаться из клетки в клетку;
|
|
- меробластическое дискоидальное дробление – характерно для рыб, птиц. При этом деление бластомеров ограничено диском цитоплазмы, свободной от желтка и расположенном на анимальном полюсе зародыша;
- мероблатстическое поверхностное (насекомые) – дробление возможно только по периферии.
Главным итогом процесса дробления является увеличение числа клеток зародыша до того критического значения, при котором в клеточных слоях начинают возникать механические напряжения, инициирующие перемещение клеток в определенные участки зародыша.
Отличие дробления от обычного митотического деления клетки заключается в следующем:
- у дробящихся бластомеров пропускается период интерфазы. Это происходит потому, что удвоение ДНК осуществляется не в S-периоде интерфазы, как при обычном митозе, а в телофазе предыдущего деления, поэтому после окончания митоза бластомер вступает в S-период интерфазы, а не в Z-период, как у соматических клеток. Таким образом, пропускается период, в котором должно произойти восстановление обычных размеров клетки, поэтому бластомеры не растут и их суммарный объем не превышает объема и массы зиготы;
- ядерно-цитоплазматическое отношение при дроблении достигает величины соматической клетки;
- в начальном периоде дробления собственный геном зародыша не активен. Усиленный синтез белка осуществляется за счет мРНК, рРНК, которые были заготовлены в яйцеклетке в период роста в овогенезе. Но на определенном этапе развития появляется потребность не только в материнской, но и в отцовской информации, и начинается трансляция с генома зародыша, причем у разных биологических видов это происходит в разные периоды эмбриогенеза: у морского ежа – на стадии тридцати двух бластомеров; у прудовика – на стадии восьми бластомеров; у млекопитающих – на стадии двух бластомеров. Такой период дробления выделяется в отдельную фазу – бластуляцию.
В процессе дробления происходит увеличение числа клеток до определенного критического уровня. Как только клетки бластулы достигают так называемой критической массы, они приобретают способность к направленным перемещениям, образуя многослойный зародыш. При этом продолжается дальнейший рост и дифференцировка клеток. Такой период эмбрионального развития, при котором клетки приобретают способность к направленным перемещениям, образуя многослойный зародыш, называется гаструляцией.
|
|
В зависимости от типа перемещения клеток и клеточных пластов условно выделяют следующие типы гаструляции.
Инвагинация – впячивание стенки бластулы (характерно для ланцетника).
Эпиболия – обрастание крупных бластомеров более мелкими делящимися бластомерами (амфибии, рыбы).
Деламинация – расщепление бластодермы на два слоя (рыбы, птицы, млекопитающие).
Иммиграция – выселение бластомеров внутрь бластулы (рыбы, птицы, млекопитающие).
Также условно выделяют раннюю гаструляцию, в ходе которой образуется двухслойный зародыш и позднюю, при которой зародыш становится трехслойным, при этом происходит закладка осевых органов – хорды, нервной и кишечной трубки.
В процессе гаструляции могут иметь место одновременно несколько типов активных перемещений бластомеров, поэтому такое разделение условно. Также условно выделяют раннюю гаструляцию, в ходе которой образуется двухслойный зародыш, и позднюю, при которой зародыш становится трехслойным. При этом происходит закладка осевых органов – хорды, нервной и кишечной трубки.
В ходе оотипической дифференцировки образуются различия между отдельными частями зиготы. Перемещения пигментных гранул и морфогенов в разные отделы зиготы активизирует определенные гены. Следовательно, первый механизм, запускающий бластомерную дифференцировку – это дифференциальная экспрессия генов. В последующем включается второй механизм – формирование межклеточных контактов между бластомерами.
Таким образом, в основе бластомерной дифференцировки лежит контактное межклеточное взаимодействие. В ходе бластомерной дифференцировки устанавливаются пути клеточных перемещений, которые осуществляются при гаструляции.
В ходе гаструляции выделяют третий этап дифференцировки – это процесс формирования отличающихся друг от друга зародышевых листков и осевых органов. В процессе гаструляции связи между клетками становятся все более тесными и появляется новый вид взаимодействия – индукция. Именно под действием индукционных контактов между клетками осуществляется этот этап дифференцировки.
|
|
Механизм клеточных перемещений у различных биологических видов различен и представляет собой сочетание тех или иных типов гаструляции – эпиболии, инвагинации, деламинации, миграции. Но независимо от того, каким типом осуществляется гаструляция, существуют общие изменения на клеточном уровне, которые приводят к клеточным перемещениям.
Это: а) клеточные деления; б) возникновение механических напряжений в пласте клеток; в) чередование растяжения и сокращения клеточного пласта. Растяжение, или поляризация клетки, осуществляется путем перераспределения органелл и вытягивание клетки в направлении предстоящего движения. Благодаря клеточным контактам процесс поляризации охватывает целый пласт клеток. Затем клетки сокращаются, что приводит к изгибанию клеточного пласта и последующему его перемещению.
В процессе ранней гаструляции образуется двухслойный зародыш, имеющий два зародышевых листка.
Зародышевый листок – это слой клеток, характеризующийся определенными тенденциями развития и принимающий участие в возникновении зачатков определенных органов, то есть каждый зародышевый листок развивается в заданном направлении.
Существует теория зародышевых листков, согласно которой:
1) зародышевые листки формируются у всех классов позвоночных;
2) один и тот же зародышевый листок у представителей различных классов позвоночных обладает одними и теми же свойствами;
3) у всех животных аналогичные органы и ткани происходят из одного и того же зародышевого листка;
4) каждый зародышевый листок представляет собой часть целого и способен дифференцироваться, только взаимодействуя между собой и находясь под воздействием влияния зародыша как целого.
Согласно теории зародышевых листков у всех классов позвоночных из одного и того же зародышевого листка формируются определенные органы и ткани, что подтверждает положение о единстве происхождения животного мира.
Так, из наружного зародышевого листка, образовавшегося на первом этапе гаструляции, в последующем формируются нервные ткани и покровный эпителий. Из внутреннего зародышевого листка – кишечный однослойный эпителий. Средний зародышевый листок – мезодерма – формируется в ходе поздней гаструляции и в последующем образует скелетные мышцы, соединительную ткань, однослойный эпителий почек, гонад и серозных полостей. В свою очередь из мезодермы и энтодермы выселяются клетки, которые заполняют пространство между зародышевыми листками и формируют мезенхиму, клетки которой образуют гладкую мускулатуру и соединительную ткань.
|
|
В процессе ранней гаструляции благодаря целенаправленным клеточным перемещениям зародыш становится двухслойным. Наружный зародышевый листок – эпибласт – у высших животных в последующем преобразуется в эктодерму; внутренний – гипобласт – в энтодерму. У низших позвоночных при этом образуется новая полость – гастроцель с отверстием – бластопором (первичный рот) и краями или губами: дорсальной, вентральной и двумя латеральными, которые соответствуют положению тела. У млекопитающих в ходе ранней гаструляции образуются аналоги бластопора и некоторые внезародышевые органы.
При поздней гаструляции формируется третий зародышевый листок – мезодерма, комплекс осевых зачатков органов (хорда, нервная и кишечная трубка) и внезародышевые органы.
Бластопор (или его аналог у позвоночных) является ориентиром для представления о клеточных перемещениях, происходящих в процессе гаструляции. Так, материал дорсальной губы бластопора в дальнейшем преобразуется в хорду, латеральный губы – в мезодерму. Судьба бластопора у различных животных различна. У первичноротых бластопор, развиваясь, превращается в дефинитивный рот; у вторичноротых – преобразуется в анальное отверстие. У высших позвоночных в процессе гаструляции бластопор вообще не образуется и можно говорить только о его аналогах.
Первой из материала дорсальной губы бластопора формируется хорда – плотный клеточный тяж, расположенный по средней линии зародыша. В дальнейшем под индукционным влиянием хорды формируется нервная трубка с последующей закладкой нервной системы. Процесс образования нервной трубки называется нейруляцией. В результате морфогенетических перемещений в ходе гаструляции группы клеток, удаленные друг от друга, сближаются настолько, что между ними становится возможен новый вид взаимодействия – индукция. Нейруляция является результатом таких взаимодействий.
Из всех зародышевых листков, но прежде всего из мезодермы, выселяется зародышевая, или вторичная, мезенхима, которая формирует ткани внутренней среды и гладкую мускулатуру. Так, эктодерма дает начало нейродермальной мезенхиме, энтодерма – энтодермальной мезенхиме. Из мезодермы выселяется дерматомная, спланхнотомная и склеротомная мезенхима. Подвижные мезенхимальные клетки мигрируют между тремя зародышевыми листками и занимают все пространство между ними. Мезенхима образована отросчатыми клетками, которые соединены друг с другом межклеточными контактами и формируют функциональный синцитий. Между клетками находится межклеточное вещество, которое образовано тонкими мезенхимальными фибриллами с тканевой жидкостью.
Функции зародышевой мезенхимы:
- является эмбриональной соединительной тканью, выполняющей в период эмбрионального развития трофическую, опорную и регуляторную функции;
- осуществляет синтез межклеточного вещества;
- является важным эмбриональным зачатком, то есть выполняет морфогенетическую функцию. По сути, это четвертый зародышевый листок, из которого образуются ткани: соединительная, гладкая мышечная, кровь плода и др.
В результате гаструляции из сферической полой бластулы образуется трехслойная зародышевая структура, у которой внутренний слой представляет собой стенку первичной кишки, сформированную энтодермой, наружный слой сформирован эктодермой и промежуточный рыхлый слой сформирован мезодермой и мезенхимой.
Эти три первичных зародышевых листка характерны для всех высших животных, формируются у всех классов позвоночных, обладают одними и теми же свойствами, что подтверждает единство происхождения животного мира.
Организация трехслойного эмбриона в общих чертах соответствует организации взрослого животного с пищеварительной трубкой внутри, эпидермисом снаружи и органами соединительнотканного происхождения между ними.
В дальнейшем энтодерма образует кишечную трубку – зачаток пищеварительной трубки, которая тянется от рта к анусу и дает начало глотке, пищеводу, желудку, кишечнику, железам. Из выростов стенки кишечной трубки в дальнейшем формируется трахея и бронхи. Сначала выросты увеличиваются в размерах, затем превращаются в систему разветвленных трубочек, которые переходят в гортань. Но энтодерма формирует только внутреннюю выстилку – эпителий внутренних органов, а поддерживающие мышечные и соединительнотканные части формирует мезодерма.
После завершения гаструляции мезодерма делится на две части – для правой и левой половины тела по бокам хорды, которая идет вдоль центральной оси тела. Хорда представляет собой клеточный тяж, над которым расположена эктодерма, а под ним – энтодерма. На определенном этапе развития клетки хорды набухают, в них появляются вакуоли, что приводит к удлинению хорды и выпрямлению зародыша. У позвоночных хорда является той осью, вокруг которой собираются клетки мезодермы, образующие в ходе органогенеза позвоночный столб. Таким образом хорда является предшественником позвоночного столба не только в процессе эволюции, но и в онтогенезе. Мезодерма дает начало соединительной ткани, сначала мезенхиме, ее клетки образуют рыхлую сеть и заполняют промежутки между другими тканями. Затем из мезодермы формируется костная, мышечная, хрящевая и фиброзная ткань, а также внутренний слой кожи – дерма. В ходе органогенеза из мезодермы формируется большая часть протоков мочеполовой системы и сердечно-сосудистой системы. Образованию соединительной ткани предшествует сомитная стадия, то есть процесс сегментации мезодермы на отдельные блоки – сомиты. Сначала участки несегментированной дорсальной мезодермы, расположенные по бокам от нервной трубки в виде единой клеточной массы, делятся на сегменты или сомиты. Сомиты обособляются последовательно один за другим по направлению от головного конца к хвостовому. Механизм образования сомитов заключается в том, что в ранней мезодерме существует механизм сопряжения клеток, который в дальнейшем исчезает, но благодаря ему клетки способны собираться в компактные группы – сомиты. Каждый сомит соответствует одному сегменту будущего скелета и в свою очередь делится на три части: склеротом, миотом и дерматом. Склеротом – часть сегмента, обращенного к хорде, в дальнейшем дает начало образованию ребер и позвонков. Дерматом – наружная часть сомита, которая прилежит к дорсальной поверхности тела зародыша и образует соединительнотканный слой кожи – дерму. Миотом представляет собой оставшуюся часть сомита, расположенную между склеротомом и дерматомом, и дает начало скелетной мускулатуре.
На стадии сомитов форма, размеры эмбриона и даже план строения у многих позвоночных приблизительно одинаковы: длина зародыша не превышает несколько миллиметров, центральная нервная система представлена нервной трубкой, кишечник и его производные заложены в виде энтодермальной трубки, соединительная ткань – в виде несегментированной мезенхимы, эпидермальный слой кожи – эктодерма, а сегментам туловища соответствуют сомиты. Но на более поздних стадиях развития эмбрионы различных биологических видов значительно отличаются друг от друга по величине, форме и дальнейшим путям развития.
Эктодерма формирует эпидермис и всю нервную систему. Процесс образования нервной системы – нейруляция – индуцируется хордой и мезодермой. По сути, нейруляция у позвоночных – это начало органогенеза. Образование нервной трубки начинается с утолщения дорсального участка эктодермы и образования нервной пластинки. Затем боковые края нервной пластинки утолщаются и образуются так называемые нервные валики. В самой пластинке появляется желобок, валики над ним сближаются, смыкаются и формируется полая трубка, которая сверху покрыта слоем эктодермы (рис. 5).

Рис. 5. Последовательные стадии (А–В) нейруляции у амфибий. Левый столбец (А–В) – сагиттальные разрезы, средний столбец (А'–В') – вид целых зародышей с дорсальной стороны, правый столбец (А"–В") – поперечные разрезы последовательных стадий: бл. – остаток бластоцеля; бп. – бластопор; мез. – мезодерма; н.в. – нервные валики; н.пл. – нервная пластинка; н.тр. – нервная трубка; п.к. – полость кишечника; пр.х. – презумптивная хорда; х – хорда; экт – эктодерма; энт – энтодерма [8].
В дальнейшем из нервной трубки формируется головной и спинной мозг. Нервная трубка отделяется от оставшейся части эктодермы и вдоль линии отделения часть клеток эктодермы поодиночке мигрирует через слой мезодермы, образуя нервный гребень. Из нервного гребня формируются все компоненты периферической нервной системы, а также клетки надпочечников, секретирующие адреналин и пигментные клетки кожи. В области головы клетки нервного гребня дифференцируются в хрящевую, костную и другие виды соединительно ткани, что составляет исключение из правил соответствия зародышевых листков трем слоям взрослого организма. Органы чувств развиваются из эктодермальных закладок: одни из нервной трубки, другие из нервного гребня и наружного слоя эктодермы.
Развитие млекопитающих
У высших (плацентарных) млекопитающих яйцеклетка содержит малое количество желтка и является вторичной олигоизолецитальной. Дробление полное, неравномерное, асинхронное. В ходе дробления наблюдается ранняя компактизация бластомеров, в результате чего образуется плотный клеточный шар – морула (рис. 11). В последующем возникает полость наполненная жидкостью – бластоцель и формируется бластоциста. В процессе образования бластоцисты бластомеры наружной клеточной массы формируют трофобласт, а бластомеры внутренней клеточной массы – эмбриобласт.
У плацентарных млекопитающих по сравнению с беспозвоночными и низшими позвоночными очень рано начинает функционировать геном зародыша. Трансляция белков с генома зародыша наблюдается со стадии восьми бластомеров.
В период дробления у млекопитающих происходит ранее обособление клеток, которые образуют внезародышевые органы, что связано с внутриутробным способом развития.

Рис. 11. Схема дробления и образования бластоцисты млекопитающих [7]: А – полное асинхронное дробление зиготы, стадии 2 и 4 бластомеров; Б – обрастание светлыми клетками темных (морула); В – образование бластоцисты, формирование полости, обособление темных клеток внутренней клеточной массы от светлых - наружной клеточной массы - трофобласта; 1 – внутренняя клеточная масса, 2 – трофобласт
Особенности развития млекопитающих:
1) в ходе развития у млекопитающих появляется необходимость в создании таких внезародышевых органов как амнион и плацента, которые защищают зародыш и обеспечивают его метаболитами матери;
2) развитие зародыша происходит в матке, что освобождает его от необходимости развиваться быстро;
3) внезародышевые органы, в частности плацента, обеспечивают зародыша питательными веществами материнского организма, поэтому яйцеклетка не нуждается в большом количестве желтка и дробление протекает с той же скоростью что и деление соматической клетки.
Гаструляция млекопитающих, как и у всех позвоночных, протекает в две фазы и заканчивается образованием трех зародышевых листков, комплекса осевых зачатков органов и внезародышевых органов.
Первая фаза гаструляции осуществляется путем деламинации и протекает одновременно с имплантацией. При этом эмбриобласт бластоцисты расщепляется на эпи- и гипобласт (рис. 10). Эпибласт является источником развития зародыша и внезародышевой амниотической эктодермы. Клетки гипобласта перемещаются по внутренней поверхности трофобласта и участвуют в образовании желточного мешка, который плотно прилегает к трофобласту. Одновременно в конце ранней гаструляции происходит формирование таких внезародышевых органов, как желточный мешок, хорион, аллантоис.
Первыми из внезародышевых органов формируются желточный мешок и аллантоис. Сначала под эпибласт выселяются клетки, которые заполняют всю полость бластоцисты и образуют первичную внезародышевую мезенхиму. В ней формируются две полости – одна над эпибластом, вторая под гипобластом. Сформировавшиеся полости обрастают клетками за счет размножения клеток обоих клеточных пластов – эпи- и гипобласта. При этом образуются два пузыря – амниотический, расположенный над будущим зародышем, и желточный – под гипобластом; материалом для пузырей станут это будущие амнион и желточный мешок. Часть мезенхимы подходит близко к трофобласту и вместе с ним формирует хорион.
К началу ранней гаструляции зародыш имеет следующее строение: снаружи находится хорион, состоящий из двух слоев – трофобласта и внезародышевой мезенхимы. Сам трофобласт разделен на два листка – симпластотрофобласт, располагающийся снаружи, и внутренний цитотрофобласт. Полость зародыша заполнена внезародышевой мезенхимой. В ней находится два пузырька – амниотический пузырек, состоящий из внезародышевой мезенхимы и внезародышевой эктодермы; и пузырек желточного мешка, сформированный внезародышевой мезенхимой и внезародышевой энтодермой. Пузырьки прилегают друг к другу и прикреплены к хориону с помощью амниотической ножки, которая образована внезародышевой мезенхимой.
Тело зародыша, образованное клетками дна амниона и крышей желточного мешка, называется зародышевым диском и состоит из эпи- и гипобласта.
Поздняя гаструляция у млекопитающих осуществляется путем миграции и частичной инвагинации (рис. 10). Основные процессы протекают в эпибласте и напоминают таковые при гаструляции у птиц с образованием первичной полоски и первичного узелка. Первичная полоска содержит материал для формирования зародышевой эктодермы, и мезодермы. Первичный узелок является источником хордального отростка. Клетки первичной полоски прорывают эпибласт, мигрируют между эпибластом и гипобластом и образуют мезодерму. С этого момента зародыш становится трехслойным.
Клетки первичного узелка мигрируют под эпибласт и образуют хордальный отросток. Часть клеток первичной полоски перемещаются по направлению к гипобласту, встраиваются в него и занимают центральное положение. Из них формируется кишечная энтодерма.
Из всех зародышевых листков (но в основном из мезодермы) выселяются клетки, которые заполняют все пространство между зародышевым листком и формируют вторичную, или зародышевую, мезенхиму.
Таким образом, ранняя гаструляция у млекопитающих, как и у всех позвоночных, заканчивается расщеплением эмбриобласта на два листка, а поздняя – образованием третьего зародышевого листка – мезодермы, комплекса осевых зачатков (хорды, нервной и кишечной трубки) и внезародышевых органов (амниона, хориона, желточного мешка).
Внезародышевые органы – временные, или провизорные, – образуются у позвоночных в процессе гаструляции и имеют характерные особенности:
- формируются вне тела зародыша;
- принимают активное участие в процессе роста и развития;
- перестают функционировать при рождении.
Впервые внезародышевые органы в процессе эволюции появились у рыб в виде желточного мешка, который выполняет трофическую функцию у всех позвоночных, кроме млекопитающих. В стенке желточного мешка формируются кровеносные сосуды, которые связывают его с сосудистой системой зародыша: по ним желток, расщепленный ферментами, поступает в организм зародыша.

Рис. 12. Схема развития внезародышевых органов у птиц [7]: 1 – скорлупа; 2 – воздушная камера; 3 – белочная оболочка (белок); 4 – амниотическая складка; 5 – зародыш; 6 – кишечная трубка; 7 – аллантоис; 8 – формирование серозной оболочки (хориона); 9 – обрастание желтка и образование желточного мешка
С выходом на сушу у позвоночных появляется необходимость еще в одном внезародышевом органе – амнионе, который дает возможность зародышу развиваться в водной среде. Необходимость водной среды для развития зародыша обусловлена тем, что она удерживает постоянную температуру окружающей среды, предохраняет зародыш от высыхания, механических повреждений, обеспечивает процессы выделения и поглощения.
Амнион (амниотическая оболочка) формируется двумя способами: при замыкании надзародышевой складки у птиц и рептилий (рис. 12) или путем раздвижения экто- и энтодермы у млекопитающих и человека (рис. 13). Независимо от способа образования зародыш оказывается окруженным амниотическими оболочками со всех сторон. Сами оболочки продуцируют жидкость, которая заполняет амниотическую полость. Химический состав амниотической жидкости изменяется по мере развития зародыша.
Функции амниотической жидкости:
- механическая защита (от повреждения, высыхания, срастания);
- выбор пути дифференцировки клеток наружной поверхности зародыша и создания необходимого давления для развития полостей – носовой, ротовой, ткани для развития легкого.
В ходе эволюции формируется орган или группа вспомогательных органов, которые обеспечивают дыхание и выделение продуктов метаболизма. Это серозная оболочка у низших и хорион, плацента и аллантоис у млекопитающих.
Серозная оболочка является самой наружной внезародышевой оболочкой, которая примыкает к скорлупе у птиц или стенке матки у млекопитающего (хорион). Эта оболочка осуществляет обмен веществ между зародышем и окружающей средой.
Аллантоис – развивается в результате выпячивания стенки задней кишки. У птиц и рептилий он выполняет дыхательную и выделительную функции. У млекопитающих проводит сосуды к зародышу и плаценте.
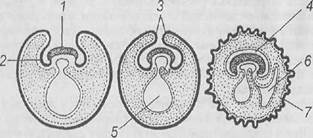
Рис. 13. Схема последовательных стадий образования внезародышевых органов у млекопитающих [7]: 1 – зародыш; 2 – туловищная складка; 3 – амниотические складки; 4 – амнион; 5 – желточный мешок; 6 – аллантоис; 7 – хорион
Хорион формируется с момента исчезновения оболочки оплодотворения и внедрения трофобласта в стенку матки, то есть с момента имплантации. В том месте, где зародыш прикрепился к слизистой оболочке матки, ворсины хориона разрастаются и вместе с децидуальной оболочкой матки формируют плаценту. На остальном протяжении они редуцируют, образуя гладкий хорион.
Первоначально ворсины представляют собой выросты трофобласта и называются первичными ворсинами. Ворсины, сформировавшиеся из трофобласта и подлежащей внезародышевой мезодерме, называются вторичными. Третичные, или истинные, ворсины состоят из эпителия трофобласта, соединительной ткани и сосудов зародыша, которые прорастают вдоль аллантоиса и разветвляются в ворсины.
Сосуды ворсин связаны с сосудами зародыша через аллантоисные сосуды, которые входят в состав пупочного канатика.
Обмен метаболитами в плаценте у млекопитающих осуществляется следующим образом: метаболиты зародыша по пупочным артериям попадают в сосуды ворсин и проникают последовательно через стенку капилляра ворсины, соединительную ткань ворсины, эпителий трофобласта и поступают в материнский организм. От матери все необходимые метаболиты проходят тот же плацентарный барьер, но уже в обратном порядке и по пупочной вене поступают к зародышу.
В зависимости от типа питания зародыша у млекопитающих выделяют следующие типы плацент (рис. 14).
1. Эпителиохориальная – ворсины хориона врастают в ткани матки и эпителий ворсин контактирует с эпителием матки.
2. Десмохориальная – ворсины хориона разрушают эпителий маточных желез и контактируют с соединительной тканью матки.
3. Эндотелиохориальная – ворсины хориона разрушают эпителий и соединительную ткань и контактируют с эндометрием сосудов.
4. Гемохориальная – ворсины хориона разрушают стенку кровеносного сосуда матки и контактируют непосредственно с кровью материнского организма.

Рис. 14. Типы плацент [2].
Животные с первым и вторым типом плацент после рождения быстро приспосабливаются к самостоятельной жизни, так как хорион только поглощает из крови матери белки и расщепляет их, а синтез собственных белков происходит в печени зародыша.
При третьем и четвертом типах плацент хорион сам синтезирует эмбриоспецифические белки из аминокислот материнского организма. Вот почему зародыши животных и человека с таким типом плацент длительное время не способны к внеутробному существованию.
КОКОРИНА Н. В.
УЧЕБНО-МЕТОДИЧЕСКОЕ ПОСОБИЕ
«ИЗБРАННЫЕ ГЛАВЫ КУРСА «БИОЛОГИЯ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ»
для студентов второго курса специальности «медицинская экология»
Минск, 2006
Автор:
Н. В. Кокорина, доцент, к.м.н.
Рекомендовано Советом факультета экологической медицины МГЭУ°им.°А.Д.°Сахарова в качестве инструктивно-методических материалов (протокол № ___ заседания Совета факультета экологической медицины от _________).
Содержание:
Лекция № 1. Молекулярно-генетические механизмы
Индивидуального развития 4
|
|
|

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...
История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

Особенности сооружения опор в сложных условиях: Сооружение ВЛ в районах с суровыми климатическими и тяжелыми геологическими условиями...

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!