КОЖНЫЕ ПОКРОВЫ И ИХ ПРОИЗВОДНЫЕ
Кожные покровы служат опорой для мышечной системы, обеспечивают регуляцию водного режима, дыхания и выделения, защищают тело от механических повреждений и проникновения вредных веществ. С кожными покровами тесно связана окраска тела насекомых. Важное значение имеют производные кожных покровов – наружные придатки (волоски, чешуйки, шипы) и внутренние, или эндоскелетные, выросты и железы (восковые, лаковые, пахучие, ядовитые и т.д.).
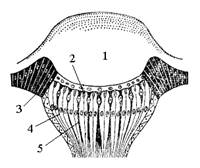
Кожные покровы состоят из трех компонентов: кутикулы, гиподермы и тонкой базальной перепонки (рис. 1).

|
| Рис. 1. Схема строения кожного покрова насекомого: 1 – гиподерма; 2 – эндокутикула; 3 – экзокутикула; 2 + 3 – прокутикула; 4 – эпикутикула; 5 – сочлененные волоски; 6 – кольцевой валик у основания волоска; 7 – шипики; 8 – поровый канал; 9 – базальная перепонка; 10 – трихогенная клетка.
|
Кутикула составляет наружную часть кожи, не имеет клеточного строения и представляет собой продукт выделения гиподермы. Образует наружный скелет насекомого, служит опорой для прикрепления мышц и, следовательно, выполняет механическую функцию. Кутикула эластична и гибка, но часто она подвергается уплотнению или склеротизации.
Кутикула подразделяется на два основных слоя – наружный и внутренний.
Наружный слой, или эпикутикула – тонкий, 1–5 мк. Эпикутикула хорошо развита у наземных насекомых, непроницаема для воды и плохо смачивается водой, т.е. гидрофобна. Непроницаемость препятствует пересыханию тела в воздушной среде. Содержит воск и липоиды, которые обеспечивают гидрофобность. Однако для ядов, растворимых в жирах, эпикутикула барьером служить не может в связи с ее липоидной структурой.
Внутренний слой, или прокутикула – в толщину несколько сот микрон, содержит 30–40% воды. Прокутикула мягкая и прозрачная, но часто наружная часть сильно склеротизуется, т.е. становится твердой и темной, образуя янтарного цвета экзокутикулу, тогда как внутренний слой сохраняет свои свойства неизменными и называется эндокутикулой. Эндокутикула прозрачна и состоит из многочисленных пластинок, имеющих волокнистое строение, что обеспечивает растяжимость кутикулы. Экзокутикула сильно склеротизована.
Биохимическую основу прокутикулы составляют хитин (25–60%) и белки. Хитин – полимерное соединение из числа азотсодержащих полисахаридов. Стоек к химическим воздействиям, нерастворим в щелочах и органических растворителях – спиртах, эфирах. Слабые кислоты на него не действуют, а сильные растворяют. Белок составляет 25–50% сухого вещества прокутикулы, может образовывать соединения с дубильными веществами. Задубленный белок плотный, темный, лишен гибкости, благодаря чему становится склеротизированным. Склеротизация образует два слоя – темный (экзокутикула) и светлый (эндокутикула). Твердость кутикулы различна: у одних насекомых (жуки) на теле образует темный панцирь, у других (личинки и имаго тлей) она очень гибкая.
Прокутикула пронизана тончайшими, менее 1 микрона поровыми канальцами, которые тянуться от гиподермы до эпикутикулы и содержат отростки плазмы гиподермальных клеток. На 1 мм тела насекомых приходятся многие тысячи каналец. Они служат путями, по которым происходит продвижение веществ из гиподермы к эпикутикуле и к поверхности экзокутикулы.
Гиподерма состоит из одного слоя клеток, образует кожный эпителий, подстилающий кутикулу. Функция гиподермы заключается в выделении и образовании кутикулы, а также в выделении линочной жидкости, которая растворяет старую эндокутикулу перед линькой насекомого.
Базальная перепонка подстилает гиподерму и не имеет клеточной структуры. Служит границей между кожными покровами и полостью тела.
Производные кожи
Придатки кожи разнообразны и подразделяются на 2 основных типа – скульптурные и структурные образования. Скульптурные – относят чисто кутикулярные образования без участия гиподермы – шипики, или хетоиды, а также бугорки, бороздки и вдавленные точки на кутикуле. Плотная поверхность кутикулы способствует развитию скульптур.
Структурные образования являются производными кожи, т.е. ее кутикулы и гиподермы. Это волоски и щетинки – хеты (chaetae). Волоски тонкие и одинаковые по всей длине. Щетинки к основанию утолщены, происходят из двух клеток гиподермы (рис. 1). Волосок выделяется тригенной клеткой, а его основание окружено мягкой перепонкой, образованной другой специализированной клеткой гиподермы. Иногда волосок вместе с перепонкой окружен валиком, образующим бугорок волоска (рис. 1), который выделяется также специальной клеткой. Если к основанию волоска подходит нервная клетка, он становится чувствительным. Производные кожи играют важную роль в регуляции температуры тела. Это чувствительные придатки, к которым присоединяется нервная клетка.
На ногах насекомых располагаются шипы и шпоры – сильные выступы, выложенные изнутри гиподермой. Они могут быть фиксированы на теле неподвижно, а могут быть сочленены подвижно. Расположение щетинок, волосков является наследственным признаком. Видоизменением хет являются чешуйки у бабочек. Чешуйки встречаются и у жуков долгоносиков и некоторых первичнобескрылых.
Эндоскелетные образования – это серия внутренних выростов кутикулы, служащих для прикрепления мышц и поддержки некоторых внутренних органов. Сильно эндоскелет развит в голове и груди, что обеспечивает прочность и фиксацию ротовых органов и крыльев. Эндоскелет головы называется тенторием.
Кожные железы могут быть одно-, двух- или многоклеточными.
Восковые железы имеются у тли, кокцид и расположены по всему телу, у пчел – на стернитах брюшка. У клопов на груди или брюшке расположены пахучие железы. Шелкоотделительные железы есть у личинок бабочек; ядовитые железы у основания волосков – у гусениц бабочек.
Окраска тела подразделяется на 2 типа – пигментную, или химическую и структурную, или оптическую. Пигментная зависит от пигмента, расположенного в кутикуле, гиподерме или в крови и жировом теле.
Основной пигмент насекомых – меланин. Это вещество белкового характера, возникающее на основе фенолов. Его цвет от желтого и светло-бурого до черного. Данное вещество представляет собой продукт обмена веществ. Меланин поглощает солнечные лучи и поэтому играет важную роль в поддержании температуры тела и изменении активности насекомых.
Желтый и красный цвет определяют каротиноиды. Их источником является каротин растений, который в мало измененном виде откладывается в коже и может находиться в крови насекомых.
Желтая и красная окраска растительноядных насекомых обеспечивается нерастворимыми в воде каротиноидами. Так, колорадский жук, питаясь листьями картофеля, поглощает каротины, которые, не перевариваясь, накапливаются в гемолимфе личинок или надкрыльях жука. К водорастворимым пигментам, также находящимся в растениях, относятся флавоноиды, которые обеспечивают желтую окраску. Зеленый цвет определяет пигмент инсектовердин.
Структурная окраска возникает вследствие особенностей строения кутикулы и расположенных на ней чешуек. Она обусловлена явлениями дифракции или интерференции, т.е. связана с особенностями разложения и отражения света. Структурная окраска свойственна жукам и чешуекрылым. Так, у бабочки медведицы на краях чешуек образуется дифракционная решетка. Видимый цвет в ней в зависимости от положения глаза наблюдателя переходит от фиолетового до красного и обратно.
Жировое тело
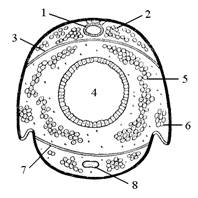
Полость тела заполнена внутренними органами и подразделена 2 тонкостенными горизонтальными перегородками – диафрагмами на 3 расположенные друг под другом отдела (рис. 2).
|
| Рис. 2. Схема поперечного разреза тела насекомого: 1 – спинной сосуд; 2 – перикардиальные клетки; 3 – верхняя диафрагма; 4 – кишечник; 5 – жировое тело; 6 – эноциты; 7 – нижняя диафрагма; 8 – брюшная нервная цепочка.
|
Верхняя диафрагма отделяет верхний, или перикардиальный, т.е. околосердечный, отдел, в нем расположен орган кровообращения – спинной сосуд. Нижняя диафрагма отделяет ниже отдел – перинейральный, т.е. околонервный. В нем расположена часть центральной нервной системы – брюшная нервная цепочка. Между верхней и нижней диафрагмами расположен средний отдел висцеральный, т.е. внутренностный. В нем находятся органы обмена – пищеварительная и выделительная системы, жировое тело, органы размножения. Дыхательная система представлена воздухоносными трубками – трахеями и трахеолами. Они пронизывают стенки всех внутренних органов и не связаны с каким-либо отделом полости тела.
Жировое тело. Это рыхлая ткань, которая пронизана трахеями и заполняет промежутки между внутренними органами. По цвету жировое тело может быть белым, желтым, оранжевым, зеленоватым. Клетки желтого тела богаты жировыми включениями и близки к гемоцитам – клеткам крови насекомых. Физиологическая роль: 1) накопление питательных веществ; 2) поглощение продуктов обмена.
Накопление питательных веществ происходит в фазе личинки, а иногда во взрослой фазе. Клетки жирового тела обогащаются жирами, углеводами, белками – гликогенами. Эти запасы расходуются во взрослом состоянии, в фазе куколки, а также в период зимовки. Поглощение продуктов обмена является уже процессом выделительной системы: клетки жирового тела накапливают соли мочевой кислоты и другие экскреты, дополняя работу мальпигиевых сосудов.
НЕРВНАЯ СИСТЕМА НАСЕКОМЫХ
Нервная система регулирует все функции организма, являясь посредником между органами чувств и другими его органами. Через органы чувств организм воспринимает информацию из внешней среды и адекватно на нее реагирует. У насекомых нервная система дифференцирована и подразделяется на центральную, периферическую и симпатическую.
Основу нервной системы составляют нервные клетки – нейроны, снабженные двумя видами отростков: 1) древовидные отростки – дендриты – короткие и сразу ветвятся на выходе из клетки; 2) аксоны – длинные, не ветвятся и только на конце есть разветвление. Нейрон имеет несколько дендритов и один аксон. Эти отростки служат для проведения нервного возбуждения и из них образуются нервы. Нервы осуществляют связь нервной системы с различными органами и частями тела (рис. 3).
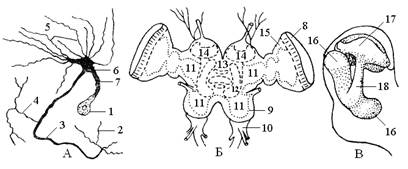
Рис. 3. Элементы нервной системы насекомых: А – нейрон; Б – головной мозг саранчи; В – схема грибовидного тела мозга таракана; 1 – нервная клетка; 2 – концевые разветвления аксона; 3 – аксон; 4 – коллатеральная ветвь аксона; 5 – дендриты; 6 – главное мозговое разветвление; 7 – клеточный отросток; 8 – протоцеребрум; 9 – дейтоцеребрум; 10 – тритоцеребрум; 11 – нейропиль; 12 – центральное тело; 13 – протоцеребральный мост; 14 – грибовидные тела; 15 – зрительная доля; 16 – лопасти ножки грибовидного тела; 17 – чашечка грибовидного тела; 18 – ножка грибовидного тела.
Различают 3 типа нейронов – чувствительные, двигательные и ассоциативные (рис. 4).
Чувствительные, или сенсорные, нейроны лежат вне центральной нервной системы, находятся на периферии тела и входят в состав органов чувств или рецепторов. Двигательные, или моторные, входят в состав нервных центров, а их аксоны заканчиваются в органах с мышцами. Реагируют на нервное возбуждение движением.
Ассоциативные нейроны входят в состав нервных центров и выполняют функцию передачи возбуждения из одного нейрона в другой, связывая чувствительные и двигательные нейроны. Передача нервного возбуждения из одного нейрона в другой или орган достигается через синапсы – это область соприкосновения отростков нейрона с другими клетками.
|
| Рис. 4. Схема взаимосвязи нейронов: А – связь нейронов с органами чувств и с органами движения; Б – схема рефлекторной дуги: 1 – нервный ганглий; 2 – дендрит; 3 – аксон; 4 – ассоативный нейрон; 5 – рецептор; 6 – чувствительный нейрон; 7 – двигательный нейрон; 8 – эффектор (мышца); 9 – окончание двигательного нерва. Стрелкой показано направление движения нервного возбуждения.
|
Центральная нервная
Система
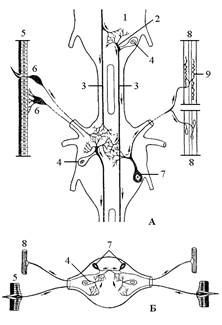
Основу центральной нервной системы составляет серия парных первичных узлов, или ганглиев, которые соединены межсегментными продольными тяжами или коннективами, а в сегменте – короткими поперечными комиссурами. Каждый сегмент тела имеет по одному парному ганглию, от которого отходят нервы, образующие периферическую нервную систему. Каждый ганглий контролирует свой сегмент тела (рис. 5). В связи с олигомеризацией (слиянием сегментов тела) это состояние утрачивается. Поэтому вся система ганглиев подразделяется на 2 отдела – головной и брюшной. Головной состоит из возвратного надглоточного узла, расположенного над кишечником, и неразвитого подглоточного узла. Они соединены тяжами, которые опоясывают передний отдел кишечника и образуют окологлоточное кольцо. Брюшной отдел состоит из серии ганглиев, расположенных под кишечником и образующих брюшную нервную цепочку.
Головной мозг в виде надглоточного узла является главным в нервной системе (рис. 6). Состоит из трех слившихся ганглиев – протоцеребрума, дейтоцеребрума и тритоцеребрума. Первый отдел – протоцеребрум развит сильнее остальных. В нем развито несколько ганглиозных центров, и наиболее сильно развита пара стебельчатых или грибовидных тел. Они являются ассоциативными и координирующими центрами нервной системы и состоят из ассоциативных нейронов. Клетки стебельчатых тел разветвлениями входят в соприкосновение с от-ростками нервных клеток и других частей нервной системы. Через эти области соприкосновения (синапсы) обеспечивается передача возбуждения и создается ассоциативная связь между клетками. С протоцеребруром связана пара зрительных долей, которая контролирует сложные глаза.
|
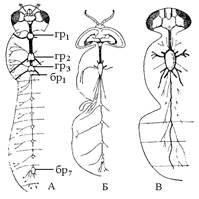
| Рис. 5. Схема разных степеней слияния узлов брюшной нервной системы у мух: А – длинноусые; Б – прямошовные короткоусые; В – круглошовные: гр – грудные узлы; бр – брюшные узлы. Цифрой обозначены порядковые номера узлов.
|
Дейтоцеребрум составляет срединный отдел головного мозга, иннервирует усики и соответствует ганглию антеннального сегмента.
Тритоцеребрум является задним отделом головного мозга. Он контролирует (иннервирует) верхнюю губу. Связан с симпатической нервной системой, так как от него начинается ее возвратный нерв.
Подглоточный узел иннервирует ротовые органы и передний отдел кишечника и является слиянием трех ганглиев челюстного отдела головы – гнатоцефалона.
Брюшная нервная цепочка состоит из трех грудных ганглиев и восьми брюшных. Последние сегменты брюшка (с 8-го по 12-й) не имеют ганглиев. Сокращение числа ганглиев в брюшной нервной цепочке достигается путем объединения как пяти брюшных, так и грудных узлов, и поэтому они объединены в 2–3 узла (может быть и в один).
Концентрация числа ганглиев в брюшной нервной цепочке – проявление олигомеризации, централизует и улучшает нервное управление организмом.
ВОЗБУЖДЕНИЕ И ТОРМОЖЕНИЕ
Возбуждение и торможение составляет основу всей нервной деятельности. Проведение возбуждения из одной части тела в другую осуществляется с помощью нервов двух типов: 1) возбуждение идет от нервной клетки к мышцам или другим органам; 2) возбуждение идет от сенсорных клеток органов чувств к нервному центру. Поэтому различают:
1) двигательные, или эфферентные, нервы – с центробежным проведением возбуждения;
2) чувствительные, или афферентные, нервы – с центростремительным проведением возбуждения.
Двигательный нерв возбуждает эффектор, чувствительный нерв получает возбуждение от рецентора. Этот путь называется рефлекторной дугой, а ответная реакция на раздражение – рефлексом (см. рис. 4).
Возбужденный нерв выделяет ацетилхолин – уксуснокислый эфир холина – азотистого вещества. Он играет роль медиатора, т.е. передатчика импульса. Для нормальной передачи возбуждения необходимо удаление избытка ацетилхолина. Удаление происходит с помощью фермента холинэстеразы, которая гидролизирует ацетилхолин на уксусную кислоту и холин.
Торможение является обратным процессом. Оно осуществляется с помощью центров торможения, которые располагаются в головном мозге или в других частях центральной нервной системы.
ОРГАНЫ ЧУВСТВ У НАСЕКОМЫХ

|
| Рис. 7. Схема кутикулярной части различных типов сенсилл. А – трихоидная сенсилла со щупальцей мухи; Б – базиконическая у саранчовых; В – стилоконическая на хоботке бабочки; Г – целоконическая у саранчовых; Д – плакоидная у наездника; Е – ампулоидная у муравья.
|
Органы чувств у насекомых являются посредниками между внешней средой и организмом. В соответствии с внешними стимулами, или раздражителями, насекомые совершают определенные действия, из которых складывается их поведение.
Органы чувств у насекомых – это механическое чувство, слух, химическое чувство, гидротермическое чувство и зрение.
Основу органов чувств составляют нервные чувствительные единицы – сенсиллы. Они состоят из двух компонентов: воспринимающей структуры в коже и прилегающих к ней нервных клеток. Сенсиллы выступают над поверхностью кожи в виде волоска, щетинки, конусы (рис. 7).
Механическое чувство. Представлено механорецепторами. Это рецепторы, а также чувствительные структуры, воспринимающие сотрясение, положение тела, его равновесие и др. Осязательные, или тактильные, рецепторы разбросаны по всему телу в виде простых сенсилл с сенсорным, т.е. чувствительным волоском. Изменение положения волоска при соприкосновениях с предметами или воздухом передается чувствительной клетке, где возникает возбуждение, передаваемое по ее отросткам в нервный центр.

|
|
Рис. 8. Схема сколопофора личинки жука-усача: 1 – концевая нить ско- лопса; 2 – колпач-ковая клетка; 3 – сколопс; 4 – обкладочная клетка; 5 – чувствительный нейрон.
|
К механорецепторам также относятся колоколовидные сенсиллы. У них отсутствуют чувствительные волоски и они погружены в кожу. Их рецепторная поверхность в виде кутикулярного колпачка находится на поверхности кутикулы. К колпачку подходит снизу стержневой отросток чувствительной клетки – штифт. Колоколовидные сенсиллы находятся на крыльях, церках, ногах, щупальцах. Они воспринимают сотрясения тела, сгибания, натяжения.
К числу механорецепторов относят и хордотональные органы как органы слуха. Их нейроны заканчиваются стержневидным штифтом. Это серия особых сенсилл, натянутых между двумя участками кутикулы. Хордотональные сенсиллы называются сколопофорами и состоят из трех клеток: чувствительного нейрона, колпачковой и обкладочной клеток.
Слух развит не у всех насекомых. У прямокрылых (кузнечики, саранчовые, сверчки), певчих цикад, некоторых клопов и ряда чешуекрылых имеются слуховые рецепторы – тимпанальные органы. Эти насекомые стрекочут или поют. Тимпанальные органы – это скопление сколопофоров, которые связаны с участками кутикулы, которые представлены в виде барабанной перепонки (рис. 8).
У саранчовых тимпанальные органы находятся по бокам 1-го сегмента брюшка, у кузнечиков и сверчков – на голенях передних ног (рис. 9).
У комаров функцию органов слуха выполняет Джонстонов орган. На церках у тараканов и прямокрылых и на теле гусениц на волосках располагаются нейроны, улавливающие звуковые волны.
Значение органов слуха:
– воспринимаются сигналы, идущие от особей своего вида, что обеспечивает связь полов, т.е. это одна из форм локации полового сигнала;
– улавливают иные звуки (свистки, резкий звук, разыскивание жертвы).

|
| Рис. 9. Передняя голень кузнечиков с отверстием тимпанального органа (ТО): 1 – окрылое овальное; 2 – щелевидное.
|
Химическое чувство. Служит для восприятия химизма среды, именно вкуса и запаха. Представлено хеморецепторами. Обоняние воспринимает и анализирует газообразную среду с низкой концентрацией вещества, а вкус – жидкую среду с высокой его концентрацией. Сенсиллы хеморецепторов представлены в виде волосков, пластинок или погруженных в тело конусов. На усиках обонятельную функцию выполняют плакоидные и целоконические сенсиллы. Обоняние служит насекомым для разыскивания особей противоположного пола, распознавания особей своего вида, для отыскания пищи и мест откладки яиц. Многие насекомые выделяют привлекающие вещества – половые аттрактанты или эпагоны.
Вкус служит только для распознавания пищи. Насекомые различают 4 основных вкуса – сладкий, горький, кислый и соленый.
Большинство сахаров, таких, как глюкоза, фруктоза, мальтоза и прочие, привлекают пчел, мух даже при сравнительно низкой концентрации; другие сахара, как галактоза, манноза и прочие, распознаются лишь при высокой концентрации, причем пчелы отвергают их. Очень чувствительны к сахарам некоторые бабочки, отличающие от чистой воды раствор сахара с ничтожной концентрацией – 0,0027%.
Многие другие вещества – кислоты, соли, аминокислоты, масла и другие – могут отвергаться при высокой концентрации, но иногда слабые растворы некоторых кислот и солей оказывают привлекающее воздействие.
Вкусовые рецепторы располагаются преимущественно на ротовых частях, но возможна и другая их локализация. Так, у пчелы, некоторых мух и ряда дневных бабочек они находятся на лапках ног и обнаруживают высокую чувствительность; при прикосновении подошвенной стороны лапок к раствору сахара голодная бабочка реагирует развертыванием хоботка. Наконец, у пчелы и складчатокрылых ос (Vespidae) эти рецепторы обнаружены и на концевых члениках усиков.
Высокая степень развития химического чувства у насекомых является существенной стороной их физиологии и служит научной основой при изыскании и применении некоторых приемов химической борьбы с вредными видами. В практике борьбы с вредителями применяют приманочный метод, сущность которого заключается в том, что те или иные привлекающие пищевые вещества обрабатываются ядами и распределяются в местах концентрации вредителя; такие отравленные приманки широко и очень успешно применяются в борьбе с саранчовыми. В борьбе с вредителями изыскиваются и привлекающие вещества, или аттрактанты.
Гигротермическое чувство. Имеет существенное значение в жизни ряда насекомых и в зависимости от условий влажности и температуры среды регулирует поведение особи; оно также контролирует водный баланс и температурный режим тела. Соответствующие рецепторы изучены недостаточно, но установлено, что ощущение влажности локализовано у некоторых насекомых на голове и ее придатках – усиках и щупальцах, ощущение тепла – на усиках, лапках и других органах. Восприятие тепла сильно развито у насекомых, и отдельные виды имеют свою оптимальную температурную зону, к которой они стремятся. Однако границы температурного оптимума зависят от условий температуры и влажности среды, в которой развивалось насекомое, а также и от фазы его развития.
Зрение. Вместе с химическим чувством, вероятно, играет решающую роль в жизни насекомых. Органы зрения имеют сложное строение и представлены двоякого рода глазами: сложными и простыми (рис. 10).

Рис. 10. Схематический разрез (А) и фасетки на поверхности (Б) сложного глаза: 1 – роговица; 2 – хрустальный конус; 3 – клетки сетчатки.
Сложные, или фасеточные, глаза в числе двух расположены по бокам головы, нередко очень сильно развиты и тогда могут занимать значительную часть головы. Каждый фасеточный глаз состоит из многозрительных единиц – сенсилл, которые называются омматидиями, число их в сложном глазу может достигать многих сотен, а также тысяч.

|
| Рис. 11. Схема строения омматидия аппозиционного глаза на продольном (А) и поперечном (Б–Г) разрезах: 1 – хрусталик; 2 – хрустальный конус; 3 – пигментные клетки; 4 – ретинальные (зрительные) клетки; 5 – рабдом.
|
Омматидий состоит из трех видов клеток, образующих соматическую, чувствительную и пигментную часть (рис. 11). Снаружи каждый омматидий образует на поверхности глаза округлую или шестигранную ячейку – фасетку, отчего сложные глаза и получили свое название. Оптическая, или преломляющая, часть омматидия состоит из прозрачного хрусталика и лежащего под ним также прозрачного хрустального конуса. Хрусталик, или роговица, является, в сущности, прозрачной кутикулой и обычно имеет вид двояковыпуклой линзы. Хрустальный конус образован четырьмя удлиненными прозрачными клетками и совместно с хрусталиком составляет единую оптическую систему – цилиндрическую линзу; длина ее оптической оси значительно превосходит ее диаметр. Чувствительная часть располагается под оптической, образует воспринимающую световые лучи сетчатку, или ретину, и состоит из серии ретинальных клеток. Эти клетки вытянуты вдоль омматидия, располагаются секториально и образуют обкладку его центрального стержня – зрительной палочки, или рабдома. У своего основания ретинальные клетки переходят в нервные волокна, идущие к зрительным долям головного мозга. Пигментная часть образована пигментными клетками, которые в совокупности составляют обкладку чувствительной части и хрустального конуса; благодаря этому каждый омматидий оптически изолирован от соседнего. Следовательно, пигментная часть выполняет функцию аппарата оптической изоляции.
Дневные насекомые имеют так называемое аппозиционное зрение. Благодаря оптической изоляции с помощью пигментных клеток каждый омматидий превращен в изолированную тонкую трубку; поэтому в него могут проникнуть только лучи, идущие через хрусталик и притом только строго совпадающие с продольной осью омматидия. Эти лучи и достигают зрительной палочки, или рабдома; последний как раз и является воспринимающим элементом сетчатки. Следовательно, поле зрения каждого омматидия очень мало и он видит только ничтожную часть рассматриваемого предмета. Но большое число омматидиев позволяет резко увеличить поле зрения путем взаимного приложения друг к другу или аппозиции; в результате из отдельных мельчайших частей изображения образуется как в мозаике единое общее изображение. Таким образом, насекомые обладают мозаичным зрением.
Ночные и сумеречные насекомые обладают суперпозиционным зрением, что связано с морфологическими и физиологическими отличиями их омматидиев. В суперпозиционном глазе чувствительная часть более отдалена от оптической, а пигментные клетки изолируют преимущественно оптическую часть. Благодаря этому к зрительной палочке проникают 2 вида лучей – прямые и косые; первые попадают в омматидий через хрусталик, а вторые – из соседних омматидиев, что усиливает световой эффект. Следовательно, изображение предмета получается в данном случае не только путем объединения отдельных восприятий, но и путем их наложения, или суперпозиции.
При сильном дневном освещении суперпозиционный глаз приобретает некоторое физиологическое сходство с аппозиционным глазом. Происходит это потому, что пигмент в пигментных клетках на свету начинает перемещаться и распределяется так, что образует темную трубку вокруг омматидия; благодаря этому омматидии оптически почти изолируются друг от друга и получают лучи преимущественно от своей линзы. Эта способность глаза реагировать на степень освещения может рассматриваться как аккомодация. В некоторой степени она свойственна и аппозиционному глазу, что позволяет дневным насекомым быстро приспособлять глаз к зрению на ярком свету и в тени, например, при перелете из открытого места в лес.
С помощью сложных глаз насекомые различают форму, движение, окраску и расстояние до предмета, а также поляризованный свет. Однако большое разнообразие насекомых, их образа жизни и повадок, несомненно, создает и разнообразие особенностей их зрения. Последние зависят от особенностей строения глаз и их омматидий; диаметр, длина, число последних и другие свойства определяют качество зрения. Считается, что многие виды близоруки и на расстоянии различают только движение. Это подтверждается многими опытами. Так, личинки стрекоз бросаются на движущуюся добычу и не замечают неподвижной. Помещенная перед гнездом ос сетка с превосходящими длину их тела ячейками все же преграждает вход в гнездо, но через некоторое время осы научатся пролезать через ячейки этой сетки.
Большинство насекомых слепы к красному цвету, но видят ультрафиолетовое излучение и привлекаются им; диапазон видимых световых волн лежит в пределах 2500–8000 А. У медоносной пчелы открыта способность различать поляризованный свет, испускаемый голубым небом, что позволяет ей ориентироваться в пространстве при полете. Для ряда насекомых характерно также изменение движения в зависимости от направления солнечных лучей, т.е. ориентация по солнечному компасу. Сущность этого явления заключается в том, что угол падения лучей на те или иные части сетчатки сохраняет свое постоянство в течение какого-то времени; прерванное движение возобновляется под тем же углом, но ввиду перемещения солнца направление движения изменяется на то же число градусов.

|
| Рис. 12. Ориентация по солнцу и искусственному источнику света. А – путь муравья до и после точки Х (в ней он был изолирован от солнца на 1 ½ ч); Б – светокомпасное движение к искусственному источнику света: α – постоянный угол.
|
Близким является светокомпасное движение, которое объясняет прилет ночных насекомых на свет. Световые лучи расходятся радиально и при косом движении по отношению к ним угол их падения будет меняться; для сохранения фиксированного угла насекомое вынуждено все время изменять свой путь в сторону источника света. Движение идет по логарифмической спирали и, в конце концов, приводит насекомое к самому источнику света (рис. 12).
Простые глаза, или глазки, располагаются между сложными глазами на лбу и темени либо только на темени (рис. 13). Они малы, обычно в числе трех, и расположены треугольником. Вследствие своего положения в верхней части головы они нередко называются также дорсальными глазками. Морфологически глазки не соответствуют омматидиям сложных глаз. Так, они иннервируются не из зрительных долей головного мозга, а из срединной части протоцеребрума. Помимо того, на одну оптическую часть у них приходится серия чувствительных частей. Они также лишены хрустального конуса и их оптическая часть представлена только кутикулярной линзой, т.е. одним хрусталиком.
|
| Рис. 13. Схема простого глаза: 1 – хрусталик; 2 – рабдом; 3 – пигментированная гиподерма; 4 – ретинальные клетки; 5 – пигментные клетки ретины.
|
Глазки развиты далеко не у всех насекомых, в частности, отсутствуют у многих двукрылых и бабочек. У бескрылых или короткокрылых насекомых они также отсутствуют или рудиментарны. Их роль недостаточно ясна. Установлено, что у ряда форм фокус глазка лежит за чувствительной частью, поэтому восприятия изображения в данном случае не может быть; закрашивание сложных глаз делает этих насекомых слепыми. Вместе с тем существует анатомическая связь глазковых нервов с нервами сложных глаз, что указывает на существование функциональной связи между этими органами. Несомненно, глазки у разных насекомых могут играть неодинаковую роль. Во всяком случае, у многих они оказывают регулирующее воздействие на сложные глаза, обеспечивая устойчивость зрения в условиях колебания интенсивности освещения. При низкой ее интенсивности глазки усиливают реакцию сложных глаз, т.е. становятся сегментами последних, при высокой – они проявляют тормозящее воздействие на сложные глаза.
От дорсальных глазков следует отличать боковые, или латеральные, глазки, свойственные личинкам насекомых с полным превращением. Эти глазки, называемые также стеммами, располагаются на боковых частях головы на месте, где у взрослых особей находятся сложные глаза. Число их различно и даже изменчиво в пределах одного и того же вида. Одни виды имеют всего лишь по одному глазку с каждой стороны, у других число их достигает шести и более пар. При переходе насекомого во взрослое состояние боковые глазки атрофируются и заменяются сложными глазами.
Стеммы разнообразны по деталям строения, но для них характерно присутствие хрусталика. У гусениц бабочек есть также хрустальный конус и развит всего один рабдом, что делает такой глазок сходным с омматидием сложного глаза. Но у личинок пилильщиков, некоторых жуков и других насекомых в глазке присутствует несколько или даже множество рабдомов, а хрустальный конус может отсутствовать. Это делает такие стеммы сходными не с омматидиями, а с дорсальными глазками.
Боковые глазки иннервируются от зрительных долей головного мозга и их зрительная функция бесспорна.
Некоторые насекомые сохраняют способность реагировать на свет при удалении глаз и глазков или покрытии их черным лаком; тараканы при этом избегают света, как и в нормальном состоянии, а гусеницы сохраняют положительную реакцию и движутся к источнику света. Безглазые пещерные насекомые также могут реагировать на свет. Очевидно, поверхность их тела способна ощущать свет и поэтому можно говорить о кожной светочувствительности.
ПОВЕДЕНИЕ НАСЕКОМЫХ
Насекомые отличаются высоко развитой способностью воспринимать и реагировать на многообразные сигналы из внешней среды – зрительные, химические, тактильные, слуховые, тепловые и др. Эти сигналы поступают из двух совершенно различных источников – от особей своего вида и от экологических факторов, но организм отвечает на них совокупностью целесообразных действий, включая и наследственно обусловленные повадки. Такая совокупная единая реакция организма обозначается понятием поведение, а раздел физиологии, изучающий его, – этологией.
Поведение определяется не только внешними раздражителями, но и зависит от внутреннего физиологического состояния особи. В его основе лежит ответная реакция на раздражение, т.е. рефлекс. В целом поведение складывается из безусловных и условных рефлексов.
Безусловные рефлексы – это врожденные реакции, т.е. рефлексы, с которыми организм рождается, наследуя их от родителей. Они составляют основу нервной деятельности насекомых, необычайно разнообразны и характерны для отдельных видов, а также родов, семейств и даже отрядов.
Безусловные рефлексы могут проявляться в виде простых актов и в виде более или менее сложных форм поведения. К числу простых актов следует отнести разнообразные движения, реакции раздражения и пр. Примером может служить рефлекс взлета у саранчи, возникающий при потере контакта ног с субстратом.
Движения и ориентацию тела по отношению к источнику раздражения часто называют таксисами или тропизмами.
Таксисы у насекомых разнообразны. К ним относятся: термотаксис – движения по отношению к источнику тепла, гигротаксис – движения по отношению к влаге, фототаксис – по отношению к свету, хемотаксис – в химическом поле раздражения, геотаксис – по отношению к земному тяготению и пр. Вследствие своего приспособительного значения таксисы могут быть положительными и отрицательными, т.е. будут направлены или к источнику раздражения, или от него. Примерами положительного таксиса могут быть случаи концентрации на






