Род Amphibacillus
Этот род описан в 1990 г. и включает один вид А. xylanus.
Прямые палочки, 0,3—0,5 х 0,9—1,9 мкм. Грамположительные на ранних стадиях роста. Эндоспоры овальные, расположенные центрально, слегка растягивающие клетки, однако быстро высвобождающиеся в результате лизиса клеток. Клетки подвижные за счет перитрихиальных жгутиков. Факультативные анаэробы; хорошо растут и спорулируют как в аэробных, так и в анаэробных условиях на средах, содержащих глюкозу, пептон и дрожжевой экстракт, при рН 10. Хемоорганотрофы. Образуют в анаэробных условиях из глюкозы этанол, уксусную и муравьиную кислоты, а в аэробных условиях — уксусную кислоту. Поглощение кислорода происходит без участия цитохромов, однако с участием HADH-оксидазы. Каталазо- и оксипазоотрицательные. Растут при температуре 25—45°С. Возможно, широко распространены в разлагающемся растительном материале.
Типовой (и единственный) вид: Amphibacillus xylanus.
Признаки вида: Разлагают ксилан и ряд других углеводов и в аэробных и в анаэробных условиях.
Выделены из навозного компоста, травы и рисовой соломы; возможно, широко распространены в навозно-травяных компостах.
Род Desulfotomaculum
Прямые или изогнутые палочки, 0,3—1,5 х 3—9 мкм, обычно расположенные одиночно. По Граму окрашиваются отрицательно, однако клеточная стенка грамположительного типа. Подвижные. Образуют овальные или сферические эндоспоры, терминальные или субтерминальные, слегка растягивающие клетки. Строгие анаэробы; хемоорганотрофы; метаболизм дыхательного типа. В качестве акцепторов электронов используют сульфат, сульфит и другие соединения серы, восстанавливая их до H2S. Содержат цитохром с протогемом в качестве простетической группы, но не цитохром с3. Каталазоотрицательные. Диапазон температуры для роста 20—70°С (оптимальный 30—55°С). Широко распространены в почве, рубце и других местообитаниях.
Типовой вид: Desulfotomaculum nigrificans.
Род Oscillospira
Крупные палочки или нити, 3—6 мкм в диаметре, разделенные близкорасположенными перегородками на многочисленные дисковидные клетки. Размножение происходит поперечным делением. Грамотрицательные; подвижные за счет многочисленных латеральных жгутиков. Могут образовывать эндоспоры. В культуре пока не получены. Инкубация на воздухе подавляет подвижность, что указывает на анаэробную природу данных организмов. Встречаются в пищеварительном тракте травоядных животных.
Типовой (и единственный) вид: Oscillospira guilliermondii.
Род Sporohalobacter
Род включает два вида: S. lortetii и S. marismortui. Палочки, 0,6—0,8 х 2,5—13 мкм. Грамотрицательные. Подвижные за счет перитрихиальных жгутиков.
Эндоспоры сферические, терминальные, растягивающие клетки в месте образования и придающие им вид барабанных палочек. Облигатные анаэробы и галофилы; для роста нуждаются в 0,5—2 М NaCl. Xe-рэорганотрофы; метаболизм бродильного типа. Обязуют ацетат, этанол, СО2 и Н2 из углеводов. Каталазо- и оксидазоотрицательные. Оптимальная температура 35—45°С. Обнаружены в соленых средах.
Типовой вид: Sporohalobacter lortetii.
Род Sporolactobacillus
Прямые палочки, 0,7—0,8 х 3—5 мкм, одиночные, в парах или, редко, в коротких цепочках. Грампложительные. Подвижные за счет перитрихи-альных жгутиков. Эндоспоры (малочисленные) овальные, терминальные, растягивающие клетки. факультативные анаэробы; дают скудный рост на воздухе. Осуществляют гомоферментативное молочнокислое сбраживание гексоз с образованием D(—)-водочной кислоты. Не содержат каталазы и цитохромов. Не восстанавливают нитрат. Не образуют индол. Оптимальная температура 35°С. Выделены из корма для цыплят и из почвы; возможно, широко распространены в природной среде.
Типовой (и единственный) вид: Sporolactobacillus mulinus.
Род Sporosarcina
Клетки сферические или овальные, 1—2 х 2—3 мкм, в основном в виде диплококков и тетрад, но иногда в виде кубических пакетов. Грамположительные. Подвижные за счет немногочисленных жгутиков, имеющихся у каждой клетки. Эндоспоры сферические (диаметр 0,5—1,5 мкм). Хемоорганотрофы. Облигатные аэробы. Хорошо растут на МПА, образуя колонии от кремовых до оранжевых. S. halophila нуждается в добавлении к среде 3% NаС1 и 0,5% MgCl2. Растут при температуре 15— 37°С. Широко распространены в почвах, включая cоленые марши.
Типовой вид: Sporosarcina ureae.
Род Sulfobacillus
Этот род описан в 1991 г.
Палочки, 0,6—0,8 х 1,0—6,0 мкм, одиночные, в парах или коротких цепочках. Грамположительные. Неподвижные. Эндоспоры сферические или слегка вытянутые, субтерминальные или терминальные, растягивающие клетки. Облигатные аэробы. Факультативные хемолитотрофы. Окисляют серу, железо и пирит, но могут адаптироваться к росту с использованием глюкозы. Оптимальный рН 1,9—2,4; оптимальная температура 50°С. Выделены из залежей пирита.
Типовой (и единственный) вид: Sulfobacillus thermosulfidooxidans.
Род Syntrophospora
Клетки палочковидные. Грамвариабельные, но клеточная стенка грамположительного типа. Эндоспоры овальные, терминальные, растягивающие клетки. Облигатные анаэробы; хемоорганотрофы. В синтрофной ассоциации с Н2-поглощающими анаэробами используют для роста насыщенные жирные кислоты, бутират и более длинноцепочечные кислоты, подвергая их р-окислению до ацетата и Н2 или, в случае нечетных неразветвленных жирных кислот, до ацетата, пропионата и Н2. Некоторые штаммы могут быть адаптированы к росту в чистой культуре за счет использования кротоната. Оптимальная температура 30°С. Сульфат и нитрат не восстанавливают. Обнаружены в пресноводных и морских осадках.
Типовой (и единственный) вид: Syntrophospora bryantii.
СПОРООБРАЗОВАНИЕ
Образование эндоспор, способность к которому в норме никогда не проявляется при вегетативном росте спорообразующих бактерий, представляет собой чрезвычайно сложный процесс дифференцировки, начинающийся во всей популяции, когда она выходит из экспоненциальной фазы роста и переходит в стационарную фазу. Этот процесс приводит к образованию внутри большинства вегетативных клеток новых клеток, полностью отличающихся от материнских по тонкой структуре, химическому составу и физиологическим свойствам. После высвобождения из материнской клетки эндоспора в норме вступает в долгий период покоя, однако при соответствующих условиях прорастает и развивается в типичную вегетативную клетку. Проблема образования и прорастания эндоспор всесторонне исследовалась многими микробиологами. Мы же ограничимся лишь кратким обзором обширной -литературы по этому вопросу. Из соображений удобства большинство экспериментальных работ по эндоспорам проведено с представителями рода Bacillus, однако образование и прорастание эндоспор у всех анаэробных спорообразующих бактерий происходит, насколько об этом можно сейчас судить, в принципе одинаковым образом.
Морфологические изменения, связанные с образованием спор обобщены на рисунке.1.
В момент перехода культур от экспоненциальной к линейной фазе роста — периоду, непосредственно предшествующему началу споруляции, — каждая клетка содержит два нуклеоида. Эти нуклеоиды объединяются, образуя палочковидную структуру. Первым видимым признаком начала спо-рообразования является возникновение поперечной перегородки около одного из полюсов клетки; эта перегородка отделяет цитоплазму и ДНК меньшей по размеру клетки, которой предстоит превратиться в спору, от остального клеточного содержимого. В отличие от того, как это бывает при обычном делении. Затем мембрана большей клетки быстро обволакивает малую, которая оказывается полностью окруженной цитоплазмой большей клетки. В результате образуется так называемая проспора. Фактически проспора представляет собой протопласт с двумя концентрическими слоями обычных мембран: собственной мембраной и мембраной материнской клетки, окружившей ее. На этой стадии процесс развития становится необратимым: можно сказать, что клетка «обязалась» осуществить споруляциях. Под световым микроскопом проспора выглядит как непреломляющая свет область, свободная от гранулярных включений.
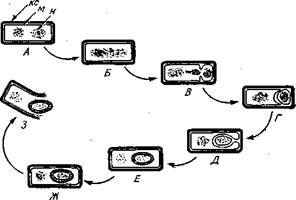
Рисунок 1. Схематическое изображение цитологических изменений, сопровождающих образование эндоспоры у Bacillus cereus.
А. Вегетативная клетка с двумя нуклеоидами (н). Б. Конденсация нуклеоидов с образованием палочковидной структуры. В. Начало формирования поперечной перегородки. Г. Завершение формирования поперечной перегородки, в результате чего проспора и ее нуклеоид оказываются изолированными от вегетативной клетки. Д. Образование новой оболочки вокруг проспоры. Е. Завершение образования оболочки. Ж. Созревание споры. 3. Высвободившаяся спора, окруженная неплотной наружной оболочкой экзоспориумом. м — клеточная мембрана, кс—клеточная стенка.
Сразу же после «поглощения» проспоры материнской клеткой происходит быстрый синтез и формирование новых структур, окружающих ее. Первым появляется кортекс, который развивается между внутренней и наружной мембранами; вскоре после этого поверх наружной мембраны начинает формироваться более электроноплотный слой, оболочка споры. У бактерий группы В. cereus вокруг оболочки образуется еще один, менее плотный и более тонкий слой, экзоспориум. После завершения формирования оболочки у созревающей споры начинает возрастать показатель преломления. В это время, однако, она еще не термостабильна. Это свойство спора приобретает вскоре после двух основных химических изменений: интенсивного поглощения спорулирующими клетками ионов Са2+ и синтеза больших количеств дипиколиновой кислоты — вещества, отсутствующего в вегетативных клетках.
Наружная оболочка споры, составляющая от 30 до 60% ее веса, состоит в основном из белков, на долю которых приходится до 80% всех белков споры. Белки оболочки обладают необычно высоким содержанием цистеина и гидрофобных аминокислот и чрезвычайно устойчивы к различным воздействиям, приводящим к растворению большинства белков.
В протопласте зрелой споры содержится очень много дипиколината кальция, а сам протопласт окружен новосинтезированными наружными слоями уникальной химической структуры (кортексом, оболочкой и иногда экзоспориумом). Высвободившаяся в результате автолиза материнской клетки эндоспора сильно обезвожена, не проявляет заметной метаболической активности и надежно защищена от тепловых и радиационных повреждений, а также от химических и ферментативных воздействий. Она остается в таком криптобиотическом состоянии до тех пор, пока ряд внешних воздействий не инициирует ее превращение в новую вегетативную клетку.
Образование эндоспоры — основное, но отнюдь не единственное изменение в спорулирующей клетке. Одно из таких примечательных явлений, свойственное Bacillus thuringensis, заключается в образовании по соседству с каждой спорой бипирамидального белкового кристаллика. Описаны также разнообразные некристаллические околоспоровые структуры определенной формы у других видов Bacillus и у Clostridium.
У многих спорообразующих бактерий, как аэробных, так и анаэробных, начало споруляции сопровождается синтезом особого класса антимикробных веществ — пептидов с мол весом около 1400. Многие из этих пептидных антибиотиков охарактеризованы и химически, и функционально. Их можно разделить на три класса: эдеины — линейные основные пептиды, подавляющие синтез ДНК, бацитрацины — циклические пептиды, подавляющие, синтез клеточных стенок, и группу грамицидин — полимиксин — тироцидин, состоящую из линейных и циклических пептидов, которые влияют на структурные и функциональные свойства мембран. Пептидные антибиотики синтезируются на относительно ранних этапах образования спор с помощью особого механизма полимеризации, при котором последовательность аминокислот определяется белком-ферментом; ни тРНК, ни рибосомы в этом процессе не участвуют. Роль указанных соединений в споруляции неизвестна, но предполагается, что они могут регулировать различные стадии процесса дифференцировки.
АКТИВАЦИЯ, ПРОРАСТАНИЕ И ДАЛЬНЕЙШЕЕ РАЗВИТИЕ СПОР
Только что сформировавшиеся эндоспоры остаются, как правило, покоящимися даже в условиях, оптимальных для прорастания. Из состояния покоя их можно вывести рядом воздействий, приводящих к так называемой активации. Вероятно, наиболее общим способом активации спор является тепловая обработка — прогрев спор в течение нескольких часов при высокой сублетальной температуре, например 65°С. Гораздо более медленная активация происходит при хранении спор как при относительно низкой температуре (5°С), так и в сухом состоянии. Эта активация необратима; по мере хранения доля спор в популяции, способных к прорастанию, возрастает.
Когда активированные споры помещают в благоприятные условия, происходит их прорастание. Процесс протекает очень быстро и проявляется в резком снижении показателя преломления спор, потере устойчивости их к нагреванию и другим вредным воздействиям, а также восстановлении метаболической активности, о чем свидетельствует внезапно возникающее дыхание. Эти процессы сопровождаются выделением в растворенном виде до 30% (по весу) веществ споры. Выделения состоят в основном из дипиколината кальция, высвобождающегося из протопластов спор, и фрагментов пептидогликана кортекса. Кортекс быстро разрушается, и остается только наружная оболочка споры. Для прорастания активированной споры необходим химический пусковой механизм; роль инициаторов этого процесса играют самые разнообразные вещества, включая L-аланин, рибозиды (аденозин, инозин), глюкозу, дипиколинат кальция и различные неорганические анионы и катионы. Специфические условия, необходимые для прорастания спор, часто меняются от вида к виду; для получения максимальной эффективности иногда необходимо сочетание нескольких веществ. Кроме того, прорастание спор можно вызвать, подвергая их физическим воздействиям, вызывающим эрозию или образование трещин в оболочке.
Процесс прорастания, по-видимому, не сопровождается синтезом макромолекул. Появление метаболической активности обусловлено активацией в протопласте споры предшествующих, но неактивных до этого ферментов. Если спора прорастает в среде, не содержащей питательных веществ, необходимых для вегетативного роста, дальнейшего развития споры не происходит; при наличии же питательных веществ, необходимых для синтеза макромолекул, прорастающая спора превращается в вегетативную клетку. При этом сначала спора набухает, а вслед за этим быстро образуется стенка вегетативной клетки, нижний слой которой, окружающий протопласт споры, может присутствовать уже во время прорастания. Новообразованная вегетативная клетка выходит из оболочки споры, удлиняется и приступает к первому вегетативному делению.





