

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Основы обеспечения единства измерений: Обеспечение единства измерений - деятельность метрологических служб, направленная на достижение...
Методика измерений сопротивления растеканию тока анодного заземления: Анодный заземлитель (анод) – проводник, погруженный в электролитическую среду (грунт, раствор электролита) и подключенный к положительному...
Техника безопасности при работе на пароконвектомате: К обслуживанию пароконвектомата допускаются лица, прошедшие технический минимум по эксплуатации оборудования...
Интересное:
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Начальные этапы развития эмбриологии
Зачатки эмбриологических знаний были уже в Древнем Египте, Вавилоне, Индии, Китае. Тем не менее, они не оказали существенного влияния на развитие представлений о становлении организмов. Первые систематические данные о развитии млекопитающих и птиц были получены учеными Древней Греции. Знаменитый врач и философ Гиппократ (460–370 гг. до н.э.) предложил двухсеменную теорию, согласно которой плод образуется через смешение мужского и женского семени (никакого представления о половых клетках в то время не существовало). Он также явился зачинателем идей преформизма, согласно которым дифференцировка частей организма происходит в некоторый начальный момент развития, а в дальнейшем наблюдается лишь их рост.
Существенный вклад в становление эмбриологии как науки внес Аристотель (384–322 гг. до н.э.), изложивший свои взгляды в сочинении «О возникновении животных». Исследования строения и развития зародышей птиц (курица), рыб (акул), моллюсков (каракатиц) и других животных позволили ему впервые сформулировать теорию эпигенеза. Согласно ей органы возникают не все сразу, а постепенно, друг за другом из вначале бесструктурной массы. Другим важным следствием его естественно-научных изысканий явилось создание учения о причинности и целесообразности, оказавшем мощное влияние на последующее развитие науки. Научная революция XVI-XVII вв. (Новое время) привела к господству механистических принципов в естествознании. В начале 17 века появились первые описания и рисунки развития куриного и человеческого зародыша (Д. Фабриций). Создание первых микроскопов способствовало накоплению огромного фактического материала в области биологии, даже с учетом известной доли бессистемности, присущей первым микроскопистам.
|
|
А. Левенгук (1632 1723) открывает сперматозоиды, Я. Сваммердам (1637 1680) проводит исследования по метаморфозу насекомых, М. Мальпиги (1628 1694) изучает развитие куриного эмбриона, работает в области микроскопической анатомии. В этот период в эмбриологии господствует теория преформизма (предвосхищения). Несовершенство микроскопов «позволяло разглядеть» зачатки всех частей будущего организма в сперматозоидах (преформисты-анималькулисты) или в яйцеклетках (преформисты-овисты). Логическим завершением данных взглядов стало формирование гипотезы «вложения», согласно которой тела потомков вложены друг в друга. Даже авторитет У. Гарвея (1578 1657), предложившего сам термин «эпигенез» и писавшего, что «ни одна часть будущего плода не существует в яйце актуально, но все части находятся в нем потенциально», не мог поколебать уверенности преформистов в своей правоте.
5) Ядро (лат. nucleus) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК), осуществляющий основные функции: хранение, передача и реализация наследственной информации с обеспечением синтеза белка. Ядро состоит из хромати́на, я́дрышка, кариопла́змы (или нуклеоплазмы) и ядерной оболочки. В клеточном ядре происходит репликация (или редуплика́ция) — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на молекуле ДНК. Синтезированные в ядре молекулы РНК модифицируются, после чего выходят в цитоплазму. Образование обеих субъединиц рибосом происходит в специальных образованиях клеточного ядра — ядрышках. Таким образом, ядро клетки является не только вместилищем генетической информации, но и местом, где этот материал функционирует и воспроизводится.
Тонкая структура клеточного ядра
Хроматин
Огромная длина молекул ДНК эукариот предопределила появление специальных механизмов хранения, репликации и реализации генетического материала. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками, необходимыми для осуществления этих процессов. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы - структуры, на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой.
|
|
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется, обычно это состояние характерно для незначимых или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфорилированием.
Считается, что в ядре существуют так называемые функциональные домены хроматина (ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной оболочки.
|
|
Ядрышко
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. N ucle o lus L ocalization S ignal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
|
|
6) Цитопла́зма (от греч. κύτος «клетка» и πλάσμα здесь «содержимое») — внутренняя среда живой или умершейклетки, кроме ядра и вакуоли, ограниченная плазматической мембраной. Включает гиалоплазму — основное прозрачное вещество цитоплазмы, находящиеся в ней обязательные клеточные компоненты — органеллы, а также различные непостоянные структуры — включения. Иногда под цитоплазмой понимают только гиалоплазму[1].
Термин «цитоплазма» ввёл Эдуард Страсбургер в 1882 году[2].
В состав цитоплазмы входят органические и неорганические вещества многих видов. Основное вещество цитоплазмы — вода. Многие вещества (например, минеральные соли, глюкоза, аминокислоты) образуют истинный раствор, некоторые другие (например, белки) — коллоидный. В ней протекают почти все процессы клеточного метаболизма. Среди прочего, в цитоплазме есть нерастворимые отходы обменных процессов и запасные питательные вещества.
Цитоплазма постоянно движется, перетекает внутри живой клетки, перемещая вместе с собой различные вещества, включения и органоиды[1]. Это движение называется циклозом.
Цитоплазма способна к росту и воспроизведению и при частичном удалении может восстановиться. Однако она нормально функционирует только в присутствии ядра. Без него долго существовать цитоплазма обычно не может[1], как и ядро без цитоплазмы.
Важнейшая роль цитоплазмы — объединение всех клеточных структур (компонентов) и обеспечение их химического взаимодействия. Она выполняет и другие функции, в частности, поддерживает тургор клетки.
Мембранные органеллы
| I. Вакуо- лярная система цито- плазмы | 1. Эндоплазма- тическая сеть | а) Другое название - эндоплазматический ретикулум. б) Это совокупность плоских мембранных мешков (цистерн), вакуолей и трубочек. |
| 2. Комплекс(аппарат)Гольджи | - Скопление 5-10 лежащих друг на друге плоских мембранных цистерн, от которых отшнуровываются мелкие пузырьки. | |
| 3. Лизосомы | - Мембранные пузырьки, содержащие ферменты гидролиза биополимеров (протеазы, нуклеазы, гликозидазы, липазы и т.д.). | |
| 4. Перокси- сомы | - Мембранные пузырьки, содержащие оксидазы - ферменты окисления субстратов непосредственно кислородом. | |
| II. Митохондрии | - Органеллы, отграниченные (как и ядро) двумя мембранами, из которых внутренняя образует многочисленные впячивания (кристы) внутрь митохондрии. |

Немембранные органеллы
|
|
| III. Рибосомы | - Небольшие частицы из двух субъединиц рибонуклеопротеидной природы. | |
| IV. Цито- скелет и его произ- водные* | 1,а. Микро- филаменты | - Нити из белка актина (d = 5 – 7 нм), пронизывающие гиалоплазму в тангенциальном направлении (вдоль длинной оси клетки или её отростка). |
| 1,б. Микро- ворсинки | - Выпячивания цитоплазмы, каркас которых составляют параллельные пучки микрофиламентов. | |
| 2. Проме- жуточные филаменты | - Нити (d = 10 нм), чей белковый состав различен в клетках разных тканей. | |
| 3,а. Микро- трубочки | - Полые трубки (d = 24 нм) из белка тубулина, имеющие, в основном, радиальную ориентацию в клетке. | |
| 3,б. Центриоли | - Пара полых цилиндров, образованных микротрубочками и лежащих возле ядра. | |
| 3,в. Реснички и жгутики | - Выпячивания цитоплазмы, каркас которых составляет аксонема - полый цилиндр из микротрубочек. |
7) Кле́точная мембра́на (или цитолемма, или плазмалемма, или плазматическая мембрана) отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды.
Основные свойства мембраны клетки:
- Проницаемость мембраны
- Полупроницаемость мембраны
- Избирательная проницаемость мембраны
- Активная проницаемость мембраны
- Управляемая проницаемость
- Фагоцитоз и пиноцитоз мембраны
- Экзоцитоз на мембране клетки
- Наличие электрических и химических потенциалов на мембране клетки
- Изменения электрического потенциала мембраны
- Раздражимость мембраны. Обусловлена она наличием на мембране специфических рецепторов, которые контактируют с сигнальными веществами. В результате этого, зачастую, меняется состояние, как самой мембраны, так и всей клетки. После соединения с лагандами (управляющими веществами), молекулярные рецепторы, расположенные на мембране, запускают биохимические процессы.
- Каталитическая ферментативная активность мембраны клетки. Ферменты действуют как снаружи мембраны клетки, так и изнутри клетки.
Строение мембраны клетки
Клеточная мембрана трехслойна. Центральный слой – жировой служит, непосредственно, для изоляции клетки. Водорастворимые вещества он не пропускает, только жирорастворимые.
Остальные же слои – нижний и верхний представляют собой белковые образования, разбросанные в виде островков на жировом слое. Между этими островками скрываются транспортёры и ионные канальцы, которые служат именно для транспорта водорастворимых веществ как в саму клетку, так и за ее пределы.
Более подробно, жировая прослойка мембраны состоит из фосфолипидов и сфинголипидов.
Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклин-зависимые киназы и циклины.Клетки, находящиеся в G0 фазе, могут вступать в клеточный цикл при действии на них факторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своими рецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции геновциклинов и циклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различныхциклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса. Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
Типы митоза
Выработка единой типологии и классификации митозов осложняется целым спектром признаков,[~ 1] которые в различных комбинациях создают разнообразие и неоднородность картин митотического деления. При этом отдельные варианты классификации, разработанные применительно к одним таксонам, являются неприемлемыми в отношении других, поскольку не учитывают специфики их митозов. Например, отдельные варианты классификации митозов, свойственных животным или растительным организмам, оказываются неприемлемыми для водорослей.
Одним из ключевых признаков, лежащих в основе различных типологий и классификаций митотического деления, является поведение ядерной оболочки. Если образование веретена и само митотическое деление протекает внутри ядра без разрушения ядерной оболочки, то такой тип митоза называют закрытым. Митоз с распадом ядерной оболочки, соответственно, называется открытым, а митоз с распадом оболочки только на полюсах веретена, с образованием «полярных окон» — полузакрытым.
Ещё одним характерным признаком является тип симметрии митотического веретена. При плевромитозе веретено деления билатерально симметрично либо асимметрично и состоит, как правило, из двух полуверетён, располагающихся в метафазе-анафазе под углом друг к другу. Для категории ортомитозов характерна биполярная симметрия веретена деления, а в метафазе зачастую наблюдается различимая экваториальная пластинка.
В рамках обозначенных признаков наиболее многочисленным является типичный открытый ортомитоз, на примере которого ниже рассматриваются принципы и стадии митотического деления. Данный тип митоза характерен для животных, высших растений и некоторых простейших.
Варианты классификации митозов
| 7 типов митоза простейших[18]: · Закрытый внутриядерный плевромитоз · Закрытый внутриядерный ортомитоз · Закрытый эвгленоидный митоз · Закрытый внеядерный плевромитоз · Полузакрытый плевромитоз · Полузакрытый ортомитоз · Открытый ортомитоз (эумитоз) | 6 типов митоза водорослей[17]: · Закрытый центрический · Закрытый ацентрический · Полузакрытый центрический · Полузакрытый ацентрический · Открытый центрический · Открытый ацентрический | 
 Основные типы митоза у водорослей: 1. Закрытый центрический; 2. Закрытый ацентрический; 3. Полузакрытый центрический; 4. Полузакрытый ацентрический; 5. Открытый центрический; 6. Открытый ацентрический
Основные типы митоза у водорослей: 1. Закрытый центрический; 2. Закрытый ацентрический; 3. Полузакрытый центрический; 4. Полузакрытый ацентрический; 5. Открытый центрический; 6. Открытый ацентрический
|
Регуляция митоза
Основными регулято́рными механизмами митоза являются процессы фосфорилирования и протеолиза[25]. Обратимые реакции фосфорилирования и дефосфорилирования обеспечивают протекание обратимых событий митоза, таких как сборка/распад веретена деления или распад/восстановление ядерной оболочки. Протеолиз лежит в основе необратимых событий митоза, таких как разделение сестринских хроматид в анаф
15) Механизм движения клетки
Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно со скоростью 1/3 от скорости жгутика и в обратном направлении, а также приобретает поступательное движение.
Подавляющее большинство наделённых жгутиком бактерий имеют палочковидную форму. Из гидродинамических расчётов следует, что для наиболее эффективного движения отношение длины клетки к ширине должно составлять 3,7. Движение кокков крайне неэффективно, поэтому они чаще всего неподвижны.
У ряда бактерий мотор и жгутик могут вращаться только в одном направлении, переориентация происходит при остановке за счёт броуновского движения. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один пучок. Для совершения кувырка они либо меняют направление вращения, либо изменяют его скорость, из-за чего пучок распадается. При полярном расположении жгутиков один из них может служить и толкающим, и тянущим приспособлением в зависимости от направления вращения.
Скорости движения бактерий варьируют от 20 мкм/с у некоторых Bacillus до 200 мкм/с у Vibrio.
16.
Сперматозо́ид (от др.-греч. σπέρμα (род. п. σπέρματος) — семя, ζωή — «жизнь» и εἴδος — «вид») — мужская половая клетка, мужская гамета, которая служит для оплодотворения женской гаметы, яйцеклетки. Термин используется для обозначения мелких, обычно подвижных гамет у организмов, которым свойственна оогамия. Обычно они значительно меньше яйцеклетки, поскольку не содержат столь значительного количества цитоплазмы и производятся организмом одновременно в значительном количестве. Понятие «сперматозоид» необходимо отличать от понятия «сперма», поскольку последняя состоит из семенной жидкости (в которой содержатся сперматозоиды), а также содержит небольшое количество эпителиальных клетокмочеиспускательного канала. Синонимы: живчик, иногда спермий. Обычно (особенно в ботанике) спермиями называют сперматозоиды, лишенные жгутиков.
Строение и функция

Строение сперматозоида
Сперматозоид человека — это специализированная клетка, строение которой позволяет ей выполнить свою функцию: преодолеть половые пути женщины и проникнуть в яйцеклетку, чтобы внести в неё генетический материал мужчины. Сперматозоид, сливаясь с яйцеклеткой, оплодотворяет её.
В организме человека сперматозоид является самой маленькой клеткой тела (если учитывать только саму головку без хвостика). Общая длина сперматозоида у человека равна приблизительно 55 мкм. Головка составляет приблизительно 5,0 мкм в длину, 3,5 мкм в ширину и 2,5 мкм в высоту, средний участок и хвостик — соответственно, приблизительно 4,5 и 45 мкм в длину.[1]
Малые размеры, вероятно, необходимы для быстрого движения сперматозоида. Для уменьшения размера сперматозоида при его созревании происходят специальные преобразования: ядро уплотняется за счет уникального механизма конденсации хроматина (из ядра удаляются гистоны, и ДНК связывается с белками-протаминами), большая часть цитоплазмы выбрасывается из сперматозоида в виде так называемой «цитоплазматической капли», остаются только самые необходимые органеллы.
Сперматозоид мужчины имеет типичное строение и состоит из головки, средней части и хвоста.
Головка сперматозоида человека имеет форму эллипсоида, сжатого с боков, с одной из сторон имеется небольшая ямка, поэтому иногда говорят о «ложковидной» форме головки сперматозоида у человека. В головке сперматозоида располагаются следующие клеточные структуры:
· Ядро, несущее одинарный набор хромосом. Такое ядро называют гаплоидным. После слияния сперматозоида и яйцеклетки (ядро которой также гаплоидно) образуется зигота — новый диплоидный организм, несущий материнские и отцовские хромосомы. При сперматогенезе (развитии сперматозоидов) образуются сперматозоиды двух типов: несущие X-хромосому и несущие Y-хромосому. При оплодотворении яйцеклетки X-несущим сперматозоидом формируется эмбрион женского пола. При оплодотворении яйцеклетки Y-несущим сперматозоидом формируется эмбрион мужского пола. Ядро сперматозоида значительно мельче ядер других клеток, это во многом связано с уникальной организацией строения хроматина сперматозоида (см. протамины). В связи с сильной конденсацией хроматин неактивен — в ядре сперматозоида не синтезируется РНК.
· Акросома — видоизмененная лизосома — мембранный пузырек, несущий литические ферменты — вещества, растворяющие оболочку яйцеклетки. Акросома занимает около половины объёма головки и по своему размеру приблизительно равна ядру. Она лежит спереди от ядра и покрывает собой половину ядра (поэтому часто акросому сравнивают с шапочкой). При контакте с яйцеклеткой акросома выбрасывает свои ферменты наружу и растворяет небольшой участок оболочки яйцеклетки, благодаря чему образуется небольшой «проход» для проникновения сперматозоида. В акросоме содержится около 15 литических ферментов, основным из который является акрозин.
· Центросома — центр организации микротрубочек, обеспечивает движение хвоста сперматозоида, а также предположительно участвует в сближении ядер зиготы и первом клеточном делении зиготы.
Позади головки располагается так называемая «средняя часть» сперматозоида. От головки среднюю часть отделяет небольшое сужение — «шейка». Позади средней части располагается хвост. Через всю среднюю часть сперматозоида проходит цитоскелет жгутика, который состоит из микротрубочек. В средней части вокруг цитоскелета жгутика располагается митохондрион — гигантская митохондрия сперматозоида. Митохондрион имеет спиральную форму и как бы обвивает цитоскелет жгутика. Митохондрион выполняет функцию синтеза АТФ и тем самым обеспечивает движение жгутика.
Хвост, или жгутик, расположен за средней частью. Он тоньше средней части и значительно длиннее её. Хвост — орган движения сперматозоида. Его строение типично для клеточных жгутиков эукариот.
17.
Яйцеклетка (науч. ооцит, реже. овоцит) — женская гамета животных, высших растений, а также многих водорослей и другихпротистов, которым свойственна оогамия. Как правило, яйцеклетки — гаплоидные клетки, но могут иметь другую плоидность уполиплоидных организмов.
Человеческая яйцеклетка имеет диаметр примерно 130 мкм и весит 1.15 нанограмм.
В цитоплазме яйцеклеток (ооплазме) содержатся совокупность питательных веществ — желток.
Яйцеклетки образуются в результате оогенеза. После оплодотворения из оплодотворенной яйцеклетки (зиготы) развиваетсяэмбрион. При партеногенезе эмбрион, а затем новый организм развивается из неоплодотворенной яйцеклетки.
Строение яйцеклетки представлено на рисунке.
1.Ядро
2.Цитолемма
3.Фолликулярный эпителий
4.Лучистый венец
5.Кортикальные гранулы
6.Желточные включения
7.Блестящая зона
8.Рецептор во фракции Zp 3-N-ацетилглюкозоамин
Яйцеклетка перемещается в ампуле реснитчатым движением клеток эпителия и ритмичной перистальтикой маточной трубы.
Яйцеклетка окружена прозрачной оболочкой (zona pellucida) и лучистым венцом (corona radiata) несет 23 хромосомы.
Особенности строения половой клетк  и
и
Строение яйцеклетки во многом обусловлено содержанием в ней компонентов, необходимых для осуществления биосинтеза белка. К ним относятся рибосомы, ферменты, т-РНК, м-РНК и их предшественники. Особые регуляторные вещества, контролирующие процессы, происходящие с самой яйцеклеткой в разный период ее жизни, играют важнейшую роль в дезинтеграции оболочки ядра. Желток яйцеклетки содержит белки, различные жиры, фосфолипиды, минеральные соли. Он обеспечивает полноценное питание зародыша. Особенности строения яйцеклетки: ее ядро содержит гаплоидный набор хромосом (23), а в ее цитоплазме находится большое количество митохондрий, элементы эндоплазматического ретикулума, свободные рибосомы, желточные включения и РНК. По периферии ядра располагаются кортикальные гранулы.
Строение яйцеклетки человек 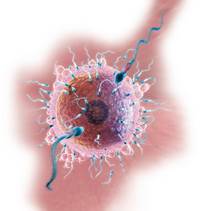 а
а
Яйцеклетка имеет специфические оболочки, выполняющие защитные функции и препятствующие проникновению в нее нескольких сперматозоидов. Они также облегчают имплантацию зародыша в стенке матки. Яйцеклетка чаще всего имеет шарообразную форму. Она содержит весь набор таких типичных органелл, какие имеет любая клетка. Яйцеклетка окружена плазматической мембраной, а снаружи покрыта блестящей оболочкой, которая состоит из мукополисахаридов. На ней находится так называемый лучистый венец или по-другому – фолликулярная оболочка, представляющая собой микроскопические ворсинки фолликулярной клетки. Она играет важнейшую защитную и питательную функцию. Только воздействие нескольких сперматозоидов способно разрушить внешнюю оболочку яйцеклетки. После такой «атаки» только один или в крайнем случае двое из них оплодотворяют ее. Яйцеклетка не имеет своего аппарата активного движения. На преодоление расстояния в 10 см, которое она проходит по яйцеводу до матки, ей требуется 4-7 дней. Яйцеклетке присуща плазматическая сегрегация, означающая то, что с момента оплодотворения до дробления яйца в ней происходит равномерное распределение цитоплазмы. Благодаря этому процессу в дальнейшем все клетки будущих тканей будут получать ее в необходимом количестве.
18. Сперматогене́з — развитие мужских половых клеток (сперматозоидов), происходящее под регулирующим воздействием гормонов. Одна из форм гаметогенеза.
Сперматозоиды развиваются из клеток-предшественников, которые проходят редукционные деления (деления мейоза) и формируют специализированные структуры (акросома, жгутик и пр.). В разных группах животных сперматогенез различается. У позвоночных животных сперматогенез проходит по следующей схеме: в эмбриогенезе первичные половые клетки — гоноциты мигрируют в зачаток гонады, где формируют популяцию клеток, называемых сперматогониями. С началом полового созреваниясперматогонии начинают активно размножаться, часть из них дифференцируется в другой клеточный тип — сперматоциты I порядка, которые вступают в мейоз и после первого деления мейоза дают популяцию клеток, называемых сперматоцитами II порядка, проходящих впоследствии второе деление мейоза и образующих сперматиды; путём ряда преобразований последние приобретают форму и структуры сперматозоида в ходе спермиогенеза.
Сперматогенез у человека

Схема поперечного сечения семенного канальца
Сперматогенез у человека в норме начинается в пубертатном периоде (около 12 лет) и продолжается до глубокой старости. Продолжительность полного сперматогенеза у мужчин составляет примерно 73—75 дней. Один цикл зародышевого эпителия составляет приблизительно 16 дней
Сперматозоиды образуются в яичках, а именно в извитых семенных канальцах. Стенка семенного канальца делится базальной мембраной на люминальную и адлюминальную стороны. На люминальной стороне расположены клетки Сертоли (сустентоциты) и предшественники половых клеток (сперматогонии, сперматоциты I и II порядков и сперматиды).
Сперматогонии, лежащие непосредственно на базальной мембране извитых семенных канальцев, проходят несколько последовательных стадий митотического деления. Общее количество сперматогоний в яичке мужчины составляет около 1 млрд. Различают две основные категории сперматогоний: А и В. Сперматогонии А, которые делятся митотически, сохраняют способность к делению и поддерживают свою популяцию. Остальные дефференцируются в сперматогоний В, которые «эвакуируются» клеточными контактами сустентоцитов (образуют под основанием половой клетки новый контакт и резорбируют старый). Сперматогония В делится митотически, дифференцируясь в сперматоцит I порядка, вступающий в мейоз.
В результате первого деления мейоза образуются две дочерние клетки сперматоциты второго порядка, каждый из которых содержит гаплоидный набор (23 у человека) d-хромосом. Вторичные сперматоциты расположены ближе к просвету канальца. Во втором делении мейоза образуются две сперматиды. Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды, каждая из которых обладает гаплоидным набором хромосом.
В ходе сложного процесса спермиогенеза сперматиды дифференцируются в зрелые сперматозоиды. Дифференцирующиеся сперматиды лежат в углублениях плазматической мембраны клеток Сертоли. При спермиогенезе комплекс Гольджи формирует акросому, содержащую протеолитические ферменты, которые при контакте с яйцеклеткой растворяют участок её блестящей оболочки (zona pellucida).
Сложный процесс сперматогенеза регулируется гонадотропными гормонами гипофиза и стероидными гормонами яичка. После полового созревания гипоталамус начинает выделять гонадотропный рилизинг-гормон, под влиянием которого гипофиз секретирует фолликулостимулирующий гормон (ФСГ), стимулирующий развитие и функционирование клетки Сертоли и лютеинизирующий гормон (ЛГ), стимулирующий клетки Лейдига к выработке тестостерона. Тестостерон оказывает воздействие на развитие клеток Сертоли, а также на предшественники половых клеток (в ассоциации с андроген-связывающим белком, выделяемым клетками Сертоли).
Секреторная активность гипофиза регулируется клетками Сертоли и клетками Лейтига. Тестостерон, выделяемый клетками Лейдига подавляет активность гипофиза к выработке ЛГ и ФСГ. Ингибин и эстрадиол, образующийся в клетках Сертоли, подавляют гипофиз к выработке ФСГ и клетки Лейдига к выработке тестостерона. Морфофункциональное состояние яичка регулируется гормонами аденогипофиза — ФСГ и ЛГ, причём уровень гормонов постоянный, имеются лишь незначительные колебания.
19. Овогенез — это процесс образования женских половых гамет, идет по той же схеме, что исперматогенез, но с некоторыми существенными отличиями.
В результате неравномерного распределения цитоплазмы как при первом, так и при втором делениях мейоза только в одной клетке оказывается большой запас питательных веществ, необходимых для развития будущего зародыша. Следовательно, образуется только одна зрелая яйцеклетка с гаплоидным набором хромосом (n) и три маленькие клеточки, которые впоследствии исчезают. При овогенезе наряду с мейозом происходит так называемое созревание яйцеклетки, во время которого значительно увеличивается ее объем.
Овогенез
(лат. ovum - яйцо + греч. genesis-зарождение, происхождение, развитие), процесс развития женских половых клеток (гамет), заканчивающийся формированием яйцеклеток.
У женщины в течение менструального цикла созревает лишь одна яйцеклетка. Процесс овогенеза имеет принципиальное сходство со сперматогенезом и также проходит через ряд стадий: размножения, роста и созревания. Яйцеклетки образуются в яичнике, развиваясь из незрелых половых клеток -овогониев, содержащих диплоидное число хромосом. Овогонии, подобно сперматогониям, претерпевают последовательные митотические деления, которые завершаются к моменту рождения плода.
Затем наступает период роста овогониев, когда их называют овоцитами I порядка. Они окружены одним слоем клеток - гранулёзной оболочкой - и образуют так называемые примордиальные фолликулы. Плод женского пола накануне рождения содержит около 2 млн. этих фолликулов, но лишь примерно 450 из них достигают стадии овоцитов II порядка и выходят из яичника в процессе овуляции.
Созревание овоцита сопровождается двумя последовательными делениями, приводящими к уменьшению числа хромосом в клетке вдвое. В результате первого деления, мейоза, образуется крупный овоцит II порядка и первое полярное тельце, а после второго деления - зрелая, способная к оплодотворению и дальнейшему развитию яйцеклетка с гаплоидным набором хромосом и второе полярное тельце. Полярные тельца, представляющие собой мелкие клетки, не играют роли в овогенезе и в конечном счёте разрушаются. В отличие от образования спермиев у мужчин, которое начинается только в период полового созревания, образование яйцеклеток у женщин начинается ещё до их рождения и завершается для каждой данной яйцеклетки только после её оплодотворения. Поэтому любые неблагоприятные факторы внешней среды, начиная со стадии внутриутробного развития девочки, могут повлечь за собой генетические аномалии у её потомства.
20.
Половой процесс, или оплодотворение, или амфимиксис (др.-греч. ἀμφι- — приставка со значением обоюдности, двойственности и μῖξις — смешение) — процесс слияния гаплоидных половых клеток, или гамет, приводящий к образованию диплоидной клетки зиготы. Не следует смешивать это понятие с половым актом (встречей половых партнёров у многоклеточных животных).
Половой процесс закономерно встречается в жизненном цикле всех организмов, у которых отмечен мейоз. Мейоз приводит к уменьшению числа хромосом в два раза (переход от диплоидного состояния к гаплоидному), половой процесс — к восстановлению числа хромосом (переход от гаплоидного состояния к диплоидному).
Различают несколько форм полового процесса:
· изогамия — гаметы не отл
|
|
|

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!