В нарушенной среде
| В нарушенной, обедненной видовым составом среде сукцессия не достигает климакса и заканчивается на тем более ранних стадиях, чем больше нарушения.
| Достижение климакса при потере одного или группы видов в результате уничтожения… не есть полное восстановление природной обстановки. Фактически это новая экосистема… К такой экосистеме надо подходить как к новому природному образованию и не переносить на нее ранее выясненные закономерности.
Н.Ф.Реймерс
|
Покрытосеменные
Покрытосеменные (Angiospermae), или цветковые (Magnoliophyta) – отдел наиболее совершенных высших растений, имеющих цветок. Ранее включались в отдел семенных растений вместе с голосеменными. В отличие от последних семязачатки цветковых заключены в завязь, образованную сросшимися плодолистиками.
Цветок является генеративным органом покрытосеменных растений. Он состоит из цветоножки и цветоложа. На последнем располагаются околоцветник (простой или двойной), андроцей (совокупность тычинок) и гинецей (совокупность плодолистиков). Каждая тычинка состоит из тонкой тычиночной нити и расширенного пыльника, в котором созревают спермии. Плодолистик цветковых растений представлен пестиком, который состоит из массивной завязи и длинного столбика, вершинная расширенная часть которого называется рыльце.
Среди покрытосеменных имеются вечнозелёные и листопадные деревья, кустарники и полукустарники, однолетние и многолетние травы. Встречаются эпифиты, использующие в качестве опоры другие растения, паразиты (например, омела), высасывающие воду и органические вещества из других растений, плотоядные растения (росянка, венерина мухоловка), ловящие и переваривающие мелких насекомых.
Покрытосеменные имеют вегетативные органы, обеспечивающие механическую опору, транспорт, фотосинтез, газообмен, а также запасание питательных веществ, и генеративные органы, участвующие в половом размножении. Внутреннее строение тканей наиболее сложно из всех растений; ситовидные элементы флоэмы окружены клетками-спутницами; почти все представители покрытосеменных имеют сосуды ксилемы.
Содержащиеся внутри пыльцевых зёрен мужские гаметы попадают на рыльце и прорастают. Гаметофиты цветковых крайне упрощены и миниатюрны, что значительно сокращает длительность цикла размножения. Образуются они в результате минимального количества митозов (трёх у женского гаметофита и двух у мужского). Одна из особенностей полового размножения – двойное оплодотворение, когда один из спермиев сливается с яйцеклеткой, образуя зиготу, а второй – с полярными ядрами, образуя эндосперм, служащий запасом питательных веществ. Семена цветковых растений заключены в плод (отсюда их второе название – покрытосеменные).
Первые цветковые растения появились в начале мелового периода около 135 миллионов лет назад (или даже в конце юрского периода). Вопрос о предке покрытосеменных в настоящее время остаётся открытым; наиболее близки к ним вымершие беннеттитовые, однако, более вероятно, что вместе с беннеттитами покрытосеменные обособились от одной из групп семенных папоротников. Первые цветковые растения были, по-видимому, вечнозелёными деревьями с примитивными цветками, лишёнными лепестков; ксилема у них всё не имела сосудов.
В середине мелового периода всего за несколько миллионов лет происходит завоевание покрытосеменными суши. Одним из важнейших условий быстрого распространения покрытосеменных была их необычайно высокая эволюционная пластичность. В результате адаптивной радиации, обусловленной экологическими и генетическими факторами (в частности, анеуполидией и полиплоидизацией), образовалось огромное количество различных видов покрытосеменных, входящих в самые разные экосистемы. К середине мелового периода образовалось большинство современных семейств. С цветковыми растениями тесно связана эволюция наземных млекопитающих, птиц и, особенно, насекомых. Последние играют исключительно важную роль в эволюции цветка, осуществляя опыление: яркая окраска, аромат, съедобная пыльца или нектар – всё это средства привлечения насекомых.
Цветковые растения распространены по всему миру, от Арктики до Антарктики. В основе их систематики лежит строение цветка и соцветия, пыльцевых зёрен, семени, анатомия ксилемы и флоэмы. Почти 250 тысяч видов покрытосеменных делятся на два класса: двудольные и однодольные, различающиеся, прежде всего, по количеству семядолей в зародышах, строению листа и цветка.
Цветковые растения являются одним из ключевых компонентов биосферы: они производят органические вещества, связывают углекислоту и выделяют в атмосферу молекулярный кислород, с них начинаются большинство пастбищных цепей питания. Многие цветковые растения используются человеком для приготовления пищи, строительства жилища, изготовления различных хозяйственных материалов, в медицинских целях.
Размножение
Гаметофит — гаплоидная многоклеточная фаза в жизненном цикле растений и водорослей, развивающаяся из спор и производящая половые клетки (гаметы).
Развивается из гаплоидных спор. На гаметофите в специальных органах гаметангиях развиваются половые клетки гаметы. Гаметангии, производящие мужские гаметы, называются антеридии, а гаметангии, производящие женские гаметы — архегонии. Оплодотворение женских гамет у наземных растений, как правило, происходит в архегонии, после чего из зиготы развивается диплоидный спорофит, который первое время зависит от гаметофита. У большинства многоклеточных водорослей оплодотворение происходит в воде (изогамия, гетерогамия и оогамия) и образовавшийся в результате оплодотворения спорофит не зависит от гаметофита. В разных группах высших растений и водорослей гаметофит развит в различной степени. У одних он существует непродолжительное время (папоротники), у других преобладает в течение всей жизни (мхи).
У высших растений всегда наблюдается правильное чередование двух многоклеточных поколений — спорофита и гаметофита. У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток. Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае (в случае так называемого нормального моноспорического женского гаметофита) из 7 клеток (содержит 8 — либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра). У папоротников, хвощей и плаунов гаметофит представляет собой небольшое, но самостоятельное растение, иногда называемое заростком. Заростки папоротников и хвощей фотосинтезирующие, однолетние. Заростки плаунов живут под землей, развиваются в течение многих лет и питаются за счет симбиоза с грибами.
У мхов гаметофит при своих небольших размерах явно доминирует над спорофитом.
Гаметы у высших растений всегда образуются в результате митоза (что принципиально отличает их от гамет животных), поскольку тело гаметофита также построено из гаплоидных клеток. Поэтому фазу гаметофита ещё называют гаметофазой, или гаплофазой. Следует отметить, что на одном гаметофите одновременно могут развиваться как мужские половые органы, так и женские. Такой гаметофит называют однодомным (обоеполым). В других случаях гаметофиты растений, относящихся к одному и тому же виду, формируют либо только мужские органы, либо только женские. Такие гаметофиты называют двудомными (раздельнополыми — мужскими и женскими). Раздельнополые гаметофиты свойственны всем семенным растениям, у которых мужские гаметофиты развиваются из микроспор, образующихся в пыльниках, а женские — из мегаспор, образующихся в семяпочках.
Органом полового размножения покрытосеменных растений является цветок. Цветок — видоизмененный, укороченный, неразветвленный побег, предназначенный для образования спор и гамет и полового процесса, завершающегося образованием семян и плода.
У цветка различают цветоножку, цветоложе, околоцветник, тычинки и пестики. У некоторых цветков отдельные части могут отсутствовать.
Цветки большинства видов растений имеют и тычинки, и пестики. Такие цветки называют обоеполыми (вишня, горох). Цветки, которые имеют только пестики, называют пестичными (женскими). Цветки, которые имеют только тычинки, называют тычиночными (мужскими). В зависимости от распределения однополых цветков на растениях различают: однодомные растения — растения, у которых на одних и тех же экземплярах располагаются и женские, и мужские цветки (огурец, кукуруза, дуб); двудомные растения — растения, у которых на одних экземплярах располагаются женские, а на других — мужские цветки (крапива двудомная, конопля, облепиха); многодомные растения — растения, у которых на одних и тех же экземплярах встречаются как обоеполые, так и однополые цветки в различных количественных соотношениях (гречиха, некоторые  виды ясеня, клена).
виды ясеня, клена).
Цветоножка — междоузлие под цветком. Цветки, лишенные цветоножки, называются сидячими (цветки в соцветии корзинка у подсолнечника, астры, одуванчика).
Цветоложе — укороченная стеблевая часть цветка. На ней располагаются все остальные части цветка.
Околоцветник — стерильная часть цветка, его покров. Околоцветник может быть простым (не дифференцированным на чашечку и венчик, образованным совокупностью однородных листочков, имеющих одинаковые размеры и окраску) и двойным (дифференцированным на чашечку и венчик, отличающиеся друг от друга размерами и окраской. Простой околоцветник может быть венчиковидным (образованным ярко окрашенными листочками) или чашечковидным (образованным зелеными листочками). Цветки, лишенные околоцветника (ива, тополь), называются голыми.
Чашечка — наружная часть двойного околоцветника, представляет собой совокупность чашелистиков — видоизмененных прицветных листьев. Обычно чашелистики имеют небольшие размеры и зеленую окраску. Они сходны с обычными листьями, но устроены проще.
Различают: раздельнолистную чашечку — чашечку, образованную свободными (несросшимися) чашелистиками (капуста, лютик); сростнолистную чашечку — чашечку, образованную частично или полностью сросшимися чашелистиками (картофель, табак, горох).
Венчик — внутренняя, обычно окрашенная часть двойного околоцветника. Представляет собой совокупность лепестков, часто имеющих яркую окраску. Количество лепестков венчика может быть различным. Лепестки могут быть более или менее одинаковыми (лютик,яблоня) либо отличаться размерами и формой (фиалка, горох). В результате венчик может быть правильным, неправильным или асимметричным. Венчик, как и чашечка, может быть раздельнолепестным и сростнолепестным. Раздельнолепестной венчик состоит из свободных, несросшихся лепестков. Сростнолепестной венчик состоит из сросшихся в той или иной степени лепестков. Главная функция венчика — привлечение опылителей.
 Андроцей — совокупность тычинок одного цветка. Количество тычинок в цветке — от одной (орхидные) до нескольких сотен (некоторые кактусы). У большинства растений тычинка состоит из тычиночной нити и пыльника. Тычиночная нить — нижняя, как правило, суженная стерильная часть тычинки. Нижний конец тычиночной нити отходит от цветоложа, а верхний конец несет пыльник. Обычно тычиночные нити тонкие, длинные, округлые в сечении. Пыльник — верхняя расширенная фертильная часть тычинки. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда (микроспорангия), в которых происходит образование микроспор, а впоследствии пылинок. Связник является продолжением тычиночной нити, через него в пыльник поступают питательные вещества.
Андроцей — совокупность тычинок одного цветка. Количество тычинок в цветке — от одной (орхидные) до нескольких сотен (некоторые кактусы). У большинства растений тычинка состоит из тычиночной нити и пыльника. Тычиночная нить — нижняя, как правило, суженная стерильная часть тычинки. Нижний конец тычиночной нити отходит от цветоложа, а верхний конец несет пыльник. Обычно тычиночные нити тонкие, длинные, округлые в сечении. Пыльник — верхняя расширенная фертильная часть тычинки. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда (микроспорангия), в которых происходит образование микроспор, а впоследствии пылинок. Связник является продолжением тычиночной нити, через него в пыльник поступают питательные вещества.
Микроспорогенез — процесс образования микроспор в микроспорангиях (гнездах пыльника). Микроспоры формируются из материнских клеток — микроспороцитов, имеющих диплоидный набор хромосом. В результате мейоза каждая материнская клетка образует четыре гаплоидных микроспоры. Микроспоры быстро обособляются друг от друга.
Микрогаметогенез — процесс образования мужских половых клеток (спермиев), происходит в пыльцевом зерне, которое является мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита происходит также в гнездах пыльников тычинок и сводится к одному митотическому делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины (наружной, толстой). Каждое пыльцевое зерно содержит две гаплоидные клетки: вегетативную и генеративную. Из генеративной (спермагенной) далее образуются два спермия. Из вегетативной (сифоногенной) впоследствии образуется пыльцевая трубка.
Гинецей — совокупность пестиков одного цветка. Обычно в пестике выделяют три части: завязь, столбик и рыльце.
Завязь — замкнутая, нижняя, полая часть пестика, несущая и защищающая семязачатки. Завязь бывает: верхняя, нижняя, полунижняя. В завязи может располагаться от одного (пшеница, вишня) до нескольких тысяч (мак) семязачатков. Стенки завязи выполняют функцию защиты семязачатков от неблагоприятных факторов среды (высыхание, колебание температур, поедание насекомыми и т.д.). Внутри завязи (в семязачатках) происходит мегаспорогенез и мегагаметогенез, они принимают участие в образовании околоплодника.
Столбик — средняя более или менее удлиненная стерильная часть пестика, отходящая обычно от верхушки завязи, соединяет завязь и рыльце.
Рыльце — верхняя расширенная часть пестика, предназначено для восприятия пыльцы. Рыльце может быть разнообразной формы (двухлопастное, звездчатое, перистое и т.д.) и размера в зависимости от особенностей опыления. При отсутствии столбика рыльце называют сидячим.

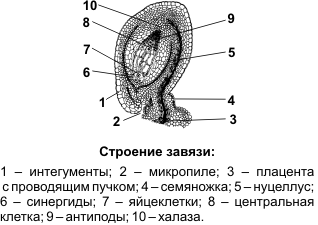
Семязачаток состоит из нуцеллуса (ядра) — центральной части, являющейся мегаспорангием, двух покровов — интегументов, которые при смыкании образуют узкий канал — микропиле, или пыльцевход, через который пыльцевая трубка проникает к зародышевому мешку. С помощью семяножки семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где сливаются нуцеллус и интегументы, называют халазой.
В семязачатке происходят мегаспорогенез, мегагаметогенез и процесс оплодотворения. После оплодотворения (реже без него) из семязачатка формируется семя.
Процесс формирования мегаспор называется мегаспорогенезом. Он происходит в нуцеллусе семязачатка. После заложения семязачатка и формирования нуцеллуса в области микропиле начинает разрастаться одна археспориальная (спорогенная) клетка — мегаспороцит, или материнская клетка мегаспор.
Материнская клетка мегаспор имеет диплоидный набор хромосом. У большинства покрытосеменных из нее путем мейоза образуются четыре гаплоидные мегаспоры. Из них лишь одна (обычно нижняя, обращенная к халазе, реже верхняя, обращенная к микропиле) дает начало женскому гаметофиту — зародышевому мешку. Остальные мегаспоры отмирают.
Процесс формирования женских половых клеток происходит в зародышевом мешке. Формирование женского гаметофита начинается с разрастания мегаспоры, которая далее три раза делится митозом. В результате этого образуются восемь клеток, которые располагаются следующим образом: три — на одном полюсе зародышевого мешка (микропилярном), три — на другом (хадазальном), две — в центре. Две оставшиеся сливаются в центре клетки, образуя диплоидную центральную клетку зародышевого мешка. Одна из трех клеток, расположенных на микропилярном полюсе, отличается большими размерами и является яйцеклеткой. Две рядом расположенные клетки являются вспомогательными и называются синергидами. Группа из трех клеток, находящихся на противоположном, халазальном полюсе, называется антиподом. Таким образом, сформированный женский гаметофит включает шесть гаплоидных клеток (яйцеклетка, две клетки-синергиды, три клетки-антипода) и одну диплоидную клетку.
Процессу оплодотворения предшествует опыление — перенос пыльцы от пыльцевых мешков тычинок к рыльцам пестиков. Попав на рыльце пестика, под воздействием веществ, выделяемых пестиком, пыльца начинает прорастать: образуется пыльцевая трубка, внедряющаяся в ткань рыльца. Кончик пыльцевой трубки выделяет вещества, размягчающие ткань рыльца и столбика. В процессе формирования пыльцевой трубки принимает участие сифоногенная клетка. По мере роста пыльцевой трубки в нее переходит спермагенная клетка, которая делится митозом с образованием двух спермиев (у некоторых растений спермагенная клетка дает начало двум спермиям еще до прорастания пыльцы). Пыльцевая трубка продвигается по столбику пестика и врастает в зародышевый мешок, как правило, через микропиле. После проникновения в зародышевый мешок кончик пыльцевой трубки разрывается, и спермии попадают внутрь. Один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу. Второй спермий сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку, из которой далее формируется эндосперм (питательная ткань) семени, обеспечивающий питание зародыша. Синергиды и антиподы дегенерируют. Вышеописанный процесс получил название двойного оплодотворения. Двойное оплодотворение у цветковых растений было открыто в 1898 году русским ботаником С.Г. Навашиным.
После двойного оплодотворения из яйцеклетки формируется зародыш семени, из центрального ядра зародышевого мешка — эндосперм, из интегументов — семенная кожура, из всего семязачатка — семя, а из стенок завязи — околоплодник. В целом из завязи пестика формируется плод с семенами.

Высшие растения
Высшие растения составляют подцарство растительного мира. Существует предположение, что они произошли от каких-то древних групп зеленых водорослей. Для такой гипотезы имеются веские основания: 1) у водорослей и высших растений главный фото-синтезирующий пигмент — хлорофилл а и каротиноиды; 2) основной запасной углевод — крахмал, который откладывается в хлоропластах, а не в цитоплазме, как у других фотосинтезирующих эукариот; 3) целлюлоза является важнейшим компонентом их клеточной стенки; 4) у водорослей и некоторых высших растений (мхи) в матриксе хлоропласта имеются особые включения — пиреноиды; 5) при клеточном делении у растений и некоторых водорослей образуются фрагмопласт — внутриклеточная пластинка, зачаток клеточной стенки.
Высшие растения появились на суше примерно 430 млн. лет назад в виде небольших по размеру и примитивных по строению риниофитов, или псилофитов. В дальнейшем их эволюция неразрывно связана с постепенным завоеванием суши. Оказавшись в совершенно иной наземновоздушной среде, они постепенно адаптировались к необычной обстановке и на протяжении многих миллионов лет дали громадное разнообразие наземных растений различной величины и сложности строения.
Одним из ключевых событий раннего этапа выхода растений на сушу было появление слор с прочными оболочками, позволяющими переносить засушливые условия и распространяться ветром. В процессе дальнейшего приспособления к наземным условиям существования у высших растений сформировались вегетативные органы — корень, стебель и лист, как результат разделения функций между различными участками тела. Корни обеспечивают закрепление растений в субстрате и водно-минеральное питание, листья — фотосинтез, стебли — транспорт веществ (восходящий и нисходящий токи).
Развитие эффективной проводящей системы тканей, состоящей из ксилемы и флоэмы, решило проблему передвижения воды и органических веществ у сосудистых растений при выходе их на сушу. От высыхания эти растения обрели защиту в виде покровной ткани — эпидермы или перидермы. Развитие эпидермы привело к возникновению устьиц — высокоспециализированных образований, регулирующих газообмен и испарениеводы растением.
Далее у высших растений произошло усиление механической прочности путем утолщения клеточной стенки и пропитывания ее лигнином, придающим жесткость ее целлюлозному остову.
Современная классификация высших растений отражает их разнообразие и историю появления на Земле: Материал с сайта http://wikiwhat.ru
- Мхи.
- Сосудистые споровые (Папоротникообразные).
- Семенные растения.
- Голосеменные.
- Покрытосеменные (Цветковые)
Параллельно шла эволюция и органов полового (антеридии и архегонии) и бесполого (спорангии) размножения. Из одноклеточных, характерных для подавляющего большинства водорослей, эти органы становятся многоклеточными, и стенки их надежно защищают развивающиеся гаметы и споры от высыхания.
В жизненном цикле высших наземных растений наблюдается закономерное чередование полового и бесполого поколений.
Гаплоидное поколение называется гаметофитом, поскольку оно способно к половому размножению и образует гаметы. Гаметы формируются в антеридиях и архегониях. В результате оплодотворения образуется зигота, из которой вырастает диплоидный спорофит. Он способен к бесполому размножению с образованием гаплоидных спор. Последние дают начало гаметофит-ному поколению. Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла высших растений.
В отличие от других высших растений, в жизненном цикле моховидных преобладает гаметофит —небольшое, преимущественно листостебельное растение, которое осуществляет функцию фотосинтеза, водоснабжения и минерального питания. Спорофит у них развивается из оплодотворенной яйцеклетки внутри архегония и постоянно связан с гаметофитом не только морфологически, но и физиологически (в смысле питания), т. е. низведен до степени органа растения, выполняющего только функцию спорообразования. Для размножения мхов необходима вода, иначе сперматозоиды не смогут подплыть к архегони-ям. Кроме того, только в достаточно влажной среде у них лопаются антеридии и высвобождаются сперматозоиды. Поэтому не случайно большинство моховидных обитает в сырых и затененных местах.
Развитие жизненного цикла моховидных по пути возрастания самостоятельности гаметофита и морфологического упрощения (с потерей самостоятельности) спорофита привело к эволюционному тупику.
В эволюции высших сосудистых растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита. Так, у плауновидных, хвощевидных ипапоротниковидных гаметофит представлен маленьким (от нескольких миллиметров до 3 см) заростком, не расчлененным на органы, живущим несколько недель (у плаунов — несколько лет) независимо от спорофита. На заростках в антеридиях развиваются сперматозоиды, которые, плавая в каплях воды, достигают архегония и сливаются с яйцеклеткой. Благодаря крошечным размерам гаметофитов оплодотворение у хвощей, плаунов и папоротников может происходить даже при ничтожно малых количествах воды в виде капелек росы, тумана и Др.
У голо- и покрытосеменных растений гаметофит полностью утратил способность к самостоятельному образу жизни, и все его развитие протекает на спорофите внутри макроспорангия (или семязачатка).
У голосеменных женский гаметофит — многоклеточный гаплоидный эндосперм с двумя (у сосны) или несколькими (у других голосеменных) архегониями; у покрытосеменных он редуцирован обычно до семи клеток, архегониев не имеет и называется зародышевым мешком. В последнем образуются яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид, вторичного диплоидного ядра и клеток-антипод.
Мужской гаметофит семенных растений развивается из микроспоры и представляет собой пыльцевое зерно (пыльцу), прорастающее в пыльцевую трубку с образованием двух спермиев. При этом впервые в эволюции растений процесс оплодотворения становится независимым от наличия капельно-жидкой среды: спермин доставляются к яйцеклеткам пыльцевой трубкой, что является важнейшим приспособлением к наземному образу жизни.
Водоросли
Водоросли — группа организмов различного происхождения, объединённых следующими признаками: наличие хлорофилла и фотоавтотрофного питания; у многоклеточных — отсутствие чёткой дифференцировки тела (называемого слоевищем, или талломом) на органы; отсутствие ярко выраженной проводящей системы; обитание в водной среде или во влажных условиях (в почве, сырых местах и т. п.). Они сами по себе не имеют органов, тканей и лишены покровной оболочки.
Некоторые водоросли способны к гетеротрофии (питанию готовой органикой), как осмотрофной (поверхностью клетки), например жгутиконосцы, так и путём заглатывания через клеточный рот (эвгленовые, динофитовые). Размеры водорослей колеблются от долей микрона (кокколитофориды и некоторые диатомеи) до 30—50 м (бурые водоросли — ламинария, макроцистис, саргассум)[1]. Таллом бывает как одноклеточным, так и многоклеточным. Среди многоклеточных водорослей наряду с крупными есть микроскопические (например, спорофит ламинариевых). Среди одноклеточных есть колониальные формы, когда отдельные клетки тесно связаны между собой (соединены через плазмодесмы или погружены в общую слизь).
К водорослям относят различное число (в зависимости от классификации) отделов эукариот, многие из которых не связаны общим происхождением. Также к водорослям часто относят сине-зелёные водоросли или цианобактерии, являющиеся прокариотами. Традиционно водоросли причисляются к растениям.
Клетки водорослей (за исключением амёбоидного типа) покрыты клеточной стенкой или клеточной оболочкой. Стенка находится снаружи мембраны клетки, обычно содержит структурный компонент (например, целлюлозу) и аморфный матрикс (например, пектиновые или агаровые вещества); также в ней могут быть дополнительные слои (например, спорополлениновый слой у хлореллы). Клеточная оболочка представляет собой или внешний кремнийорганический панцирь (у диатомей и некоторых других охрофитовых), или уплотнённый верхний слой цитоплазмы (плазмалемму), в котором могут быть дополнительные структуры, например, пузырьки, пустые или с целлюлозными пластинками (своеобразный панцирь, тека, у динофлагеллятов). Если клеточная оболочка пластичная, клетка может быть способна к так называемому метаболическому движению — скольжению за счёт небольшого изменения формы тела.
Фотосинтезирующие (и «маскирующие» их) пигменты находятся в особых пластидах — хлоропластах. Хлоропласт имеет две (красные, зелёные, харовые водоросли), три (эвглены, динофлагелляты) или четыре (охрофитовые водоросли) мембраны. Также он имеет собственный сильно редуцированный генетический аппарат, что позволяет предположить его симбиогенез (происхождение от захваченной прокариотной или, у гетероконтных водорослей, эукариотной клетки). Внутренняя мембрана выпячивается внутрь, образуя складки — тилакоиды, собранные в стопки — граны: монотилакоидные у красных и сине-зелёных, двух- и больше у зелёных и харовых, трёхтилакоидные у остальных. На тилакоидах, собственно, и расположены пигменты. Хлоропласты у водорослей имеют различную форму (мелкие дисковидные, спиралевидные, чашевидные, звёздчатые и т. д.).
У многих в хлоропласте имеются плотные образования — пиреноиды.
Продукты фотосинтеза, в данный момент излишние, сохраняются в форме различных запасных веществ: крахмала, гликогена, других полисахаридов, липидов. Помимо прочего липиды, будучи легче воды, позволяют держаться на плаву планктонным диатомовым с их тяжёлым панцирем. В некоторых водорослях образуются газовые пузыри, также обеспечивающие водоросли подъёмную силу.
У водорослей выделяют несколько основных типов организации таллома:
· Амёбоидный (ризоподиальный)
Одноклеточные организмы, лишённые твёрдой клеточной оболочки, и вследствие этого, не способные сохранять постоянную форму тела. Благодаря отсутствию клеточной стенки и наличию особых внутриклеточных структур клетка способна к ползающему движению посредством псевдоподий или ризоподий. Для некоторых видов характерно образование многоядерного плазмодия путём слияния нескольких амёбоидных клеток. Амёбоидное строение могут вторично приобретать некоторые монадные формы путём отбрасывания или втягивания жгутиков.
· Монадный
Одноклеточные водоросли, имеющие постоянную форму тела, жгутик(и), часто стигму, а пресноводные — сократительную вакуоль. Клетки активно двигаются в вегетативном состоянии. Часто встречается объединение нескольких монадных клеток в колонию, окружённую общей слизью, в некоторых случаях они даже соединяются между собой посредством плазмодесм. У высокоорганизованных форм с многоклеточным талломом часто имеются расселительные стадии — зооспоры и гаметы, имеющие монадную структуру.
· Коккоидный
Одноклеточные, лишённые каких-либо органоидов передвижения и сохраняющие постоянную форму тела в вегетативном состоянии клетки. Чаще всего имеется утолщённая клеточная стенка или панцирь, могут быть различные выросты, поры и пр. для облегчения парения в толще воды. Многим водорослям с данной структурой свойственно образование колоний. Некоторые диатомеи и десмидиевые способны к активному передвижению путём выделения слизи.
· Пальмеллоидный (капсальный)
Постоянное, достаточно крупное, как правило, прикреплённое к субстрату, образование из нескольких коккоидных клеток, погружённых в общую слизистую массу. Клетки непосредственно между собой не объединяются — отсутствуют плазмодесмы. Временную стадию жизненного цикла с аналогичной морфологией называют пальмеллевидным состоянием. В такое состояние могут переходить многие монадные и коккоидные водоросли при наступлении неблагоприятных условий, образующиеся при этом пальмеллевидные образования, как правило, мелкие и не имеют постоянной формы.
· Нитчатый (трихальный)
Клетки соединены в нить, простую или разветвлённую. Нити могут свободно плавать в толще воды, прикрепляться к субстрату, либо объединяться в колонию. Вегетативно нитчатые водоросли размножаются обычно распадом нити на отдельные фрагменты. Рост нитей может идти четырьмя путями: диффузным — делятся все клетки нити, интеркалярным — зона роста расположена в середине нити, апикальным — делением конечных клеток, и базальным — делением клеток у основания таллома. Клетки в нити не имеют жгутиков и могут быть связаны между собой плазмодесмами.
· Разнонитчатый (гетеротрихальный)
Есть две системы нитей: стелющиеся по субстрату горизонтальные и отходящие от них вертикальные. Горизонтальные нити тесно смыкаются, либо могут сливаться в псевдопаренхиматозную пластинку и выполняют, в основном, опорную функцию и функцию вегетативного размножения, вертикальные нити — преимущественно ассимиляторную функцию. Иногда может наблюдаться редукция, либо чрезмерное развитие тех или иных нитей, приводящее к вторичной утрате или нарушению характерных черт гетеротрихального строения (при редукции вертикальных нитей, например, таллом может представлять собой простую однослойную пластинку, полностью прикреплённую к субстрату).
· Пластинчатый
Многоклеточные талломы в форме пластинок из одного, двух или нескольких слоёв клеток. Возникают при продольном делении клеток, составляющих нить. Число слоёв зависит от характера образования перегородок при делении клеток. Иногда слои могут расходиться, и таллом тогда приобретает трубчатую форму (полый внутри), стенки при этом становятся однослойными.
· Сифональный (неклеточный, сифоновый)
Отсутствуют клеточные перегородки, в результате чего таллом, часто крупный и внешне дифференцированный, формально представляет собой одну клетку с большим количеством ядер.
· Сифонокладальный
Таллом представлен многоядерными клетками, соединёнными в нитчатые или иной формы многоклеточные талломы (Siphonocladales).
· Харофитный (членисто-мутовчатый)
Свойственна только харовым водорослям. Таллом крупный, многоклеточный, состоит из главного побега с ветвями и отходящими от него, иногда ветвящимися, членистыми боковыми побегами. Боковые побеги отходят от главного в области узлов, часть побега между узлами состоит, как правило, из одной крупной клетки и называется междоузлием.
· Сарциноидный
Колонии, представляющие собой группы (пачки или нитеобразные образования), которые возникают в результате деления одной исходной клетки и заключены в растягивающуюся оболочку этой клетки.
· Псевдопаренхиматозный (ложнотканевый)
Представлен слоевищами, которые образовались в результате срастания разветвлённых нитей, нередко сопровождаемого морфофункциональной дифференциацией получающихся ложных тканей.
У части сине-зелёных, зелёных и красных водорослей в слоевище откладываются соединения кальция, и оно становится твёрдым. Водоросли лишены корней и поглощают нужные им вещества из воды всей поверхностью. Крупные донные водоросли имеют органы прикрепления — подошву (уплощённое расширение в основании) или ризоиды (разветвлённые выросты). У некоторых водорослей побеги стелются по дну и дают новые слоевища.
Вегетативное размножение может осуществляться как простым разделением многоклеточного организма (фрагменты нитей Spirogyra (спирогира) или колонии (Synura) на несколько частей, так и при помощи специализированных органов. Например, бурые водоросли из порядка сфацеляриевых имеют для этой цели специальные выводковые веточки, а водоросли из порядка харовых — клубеньки на ризоидах.
Бесполое размножение водорослей осуществляется при помощи подвижных зооспор или неподвижных апланоспор. При этом протопласт клетки-спорангия делится на части и продукты деления выходят из её оболочки. Особь, на которой формируются спорангии, называют спорофитом. Часто подвижные или неподвижные споры имеют специальные названия. Например, апланоспоры, приобретающие форму материнской клетки, находясь внутри неё, называются автоспорами, а если апланоспора одевается толстой оболочкой и впадает в состояние покоя, то это гипноспора; зооспоры, утратившие жгутики, но сохранившие сократительные вакуоли и глазок, будут называться гемизооспорами.
Половое размножение. При половом размножении происходит попарное слияние гаплоидных клеток с образованием диплоидной зиготы. У водорослей есть несколько вариантов полового процесса: изогамия — слияние двух одинаковых по форме и размеру подвижных гамет; гетерогамия — слияние двух подвижных гамет одинаковых по форме, но разных по размеру; оогамия — слияние крупной неподвижной яйцеклетки с мелким подвижным сперматозоидом. Клетки, в которых образуются гаметы, называются гаметангии, а сами растения с гаметангиями — гаметофитами. Гаметангии бывают двух типов: образующие многочисленные сперматозоиды — антеридии и образующие одну или несколько яйцеклеток оогонии. Также половое размножение в некоторых группах водорослей может осуществляться и без образования гамет. Например, если сливаются две подвижные одноклеточные водоросли, то это хологамия; слияние протопластов двух неподвижных гаплоидных вегетативных клеток с образованием зиготы — конъюгация. Если гаметофиты и спорофиты в жизненном цикле водоросли имеют выраженные морфологические различия, то это называется гетероморфная смена форм р





