Лекция № 5
1. Отдел Папоротниковидные – Polypodiophyta
2. Группа Праголосеменные – Progymnospermae
3. Класс Ужовниковые – Ophioglossopsida
4. Класс Полиподиевые – Polypodiopsida
Рис. 2. Схема строения листа папоротника
1 – черешок; 2 – пластинка листа; 3 – перо первого порядка; 4 – перышки (или перья второго порядка); 5 – лопасть перышка; 6 – рахис.
Крупные и сложные листовые следы и наличие листовых прорывов в стеле обнаруживаются у большинства видов. Листья папоротников отличаются большим разнообразием. В большинстве случаев листья совмещают две функции – фотосинтез и спороношение. У многих (Matteuccia struthiopteris, Onoclea sensibilis, Drynaria) листья дифференцированы на стерильные и фертильные. Считается, что все 3 типа листьев существовали в раннем палеозое независимо.
У большинства представителей семейства Polypodiaceae черешки соединяются с корневищем посредством особого сочленения или без него (возможность свободно менять ориентацию к источнику света), наличие сочленения является систематическим признаком. Для систематики имеет значение также строение проводящей системы черешка, в частности форма проводящего пучка на поперечном срезе (у осмундовых С-образная форма, у Plagiogyriaceae – Y-образная форма).
Важное значение имеет число проводящих пучков – 1,2 или несколько. У Aspleniaceae обычно 3-7, но у большинства только 2, соединенные наверху в один.
Примитивные листья имеют дихотомическое ветвление (равно- или неравнодихотомическое). У подавляющего большинства листья перистые – однажды, дважды или многократно. Пластинка перистого листа в отличие от дихотомического имеет стержень или рахис (от греч. rhachis – «позвоночник»), представляющий собой продолжение черешка. Стержень соответствует главной жилке целого листа, однорядные сегменты называются перьями (могут быть цельными, лопастными). У папоротников дихотомическое жилкование встречается у примитивных форм – открытое, не образующее сети (представитель семейства Гименофилловых – Trichomanes reniforme). Для более продвинутых форм характерно сетчатое жилкование, которое возникло не сразу.
У Папоротниковидных встречаются почти все основные типы устьичного аппарата (за исключением анизоцитного) – десмоцитный, перицитный, полоцитный. Тип устьичного аппарата специфичен и является систематическим признаком.
Размножение папоротников осуществляется в основном за счет спор. Большинство видов – равноспоровые растения. Для всех папоротников характерно отсутствие специализированных спороносных побегов – стробилов. В основном спорангии сгруппированы в сорусы; у мараттиевых сорусы, срастаясь между собой, образуют синангии. Если у примитивных видов одиночные спорангии располагаются по краям листьев либо на вершинах лопастей, при этом каждый спорангий снабжен самостоятельной жилкой, то у большинства папоротников спорангии или сорусы располагаются на нижней поверхности листьев. Место прикрепления спорангия к листу называется плацентой (ложе). В ходе эволюции происходило разрастание плаценты, и она приобретала продолговатую или шаровидную форму. Кроме этого, спорангии на плаценте располагаются в определенной последовательности, следовательно, созревание и вскрывание неодновременное. Увеличение длины ножки увеличивает количество спорангиев на плаценте. Эти два процесса – разрастание плаценты и увеличение длины ножки спорангия – происходили независимо в разных систематических группах. Остановимся подробнее на строении спорангиев, размещение которых является диагностическим признаком.
Листья папоротников возникли в процессе эволюции из ветвей риниофитовых предков. Очевидно, что спорангии древнейших Папоротниковидных имели верхушечное (терминальное) расположение, т.е. сидели на верхушках конечных веточек дихотомически разветвленного листа. Среди ныне живущих наиболее близкое к верхушечному расположению спорангиев наблюдается у гроздовника (Botrychium), относящегося к примитивным Ужовниковым. Следующий тип расположения спорангиев – краевое (маргинальное), наблюдаемый у многих папоротников, можно рассматривать как результат возникновения листовой пластинки.
Значительным шагом вперед в приспособительной эволюции папоротников был переход на нижнюю сторону листовой пластинки. Наряду с перемещением спорангиев на нижнюю сторону пластинки происходит и другое изменение. Сначала спорангии бывают расположены одиночно (у Схизейных не образуют групп). Но уже у некоторых папоротников с краевым расположением (Dicksonia, Loxsoma) или у всех Гименофилловых спорангии сгруппированы в отдельные спорангиальные кусочки, или сорусы (от греч. soros – «куча»), что дает возможность концентрированного, более эффективного снабжения питательными веществами (рис. 3).

Рис. 3. Расположение спорангиев и сорусов папоротников
1 – спорангии орляка (Pteridium); 2 – сорусы вудсии (Woodsia); 3 – сорусы с покрывальцем щитовника (Dryopteris); 4 – сорусы, встречающиеся у родов Asplenium, Athyrium, Polypodium.
В биологическом отношении чрезвычайно важен порядок развития и созревания спорангиев на листе или в каждом отдельном сорусе. У вымерших Папоротниковидных, и ныне живущих Ужовниковых, Мараттиевых, Осмундовых, Плагиогириевых, Схизейных, Глейхениевых и Матониевых все спорангии развиваются одновременно (внутри соруса), т.е. симультанно (представители семейств Плагиогириевых, Глейхениевых и Матониевых). Сорусы, в которых происходит одновременное развитие и созревание спорангиев, называют простыми. Их существенный недостаток – при ухудшении внешних условий подвергается риску и угнетению произрастание и развитие спор.
Выход – в последовательном развитии и созревании спорангиев в сорусе. Это достигается посредством удлинения плаценты (рецептакула) соруса и увеличения числа спорангиев при уменьшении их размеров.
Устанавливается базипетальная (от греч. basis – «основание», от лат. petere – «устремляться») последовательность, когда развитие начинается с верхней части плаценты (в частности, представители семейства гименофилловых имеют длинное, цилиндрическое ложе). Сорусы с базипетальным развитием называют градатными (от лат. gradatio – «постепенное возвышение, усиление»). Такие сорусы встречаются у Циатейных, Асплениевых, у всех разноспоровых. Смешанные сорусы возникают независимо в разных линиях эволюции.
В дальнейшем произошло важное событие – появились защитные приспособления. Так, в простейших случаях сорусы прикрываются загибающимся краем листа, у более специализированных видов формируется индузий (от лат. indusium – «верхняя туника»). У большинства семейств с простыми сорусами индузий отсутствует.
Другая форма защиты – это срастание в синангии (у некоторых Мараттиевых). Исключение – род Matonia, у которого настоящее покрывало встречается с простыми сорусами.
Индузий имеют папоротники с градатными сорусами. По характеру заложения и строению стенки спорангия папоротники подразделяются на лепто - и эуспорангиатные.
У эуспорангиатных папоротников спорангий возникает из группы клеток и имеет многослойную стенку, у лептоспорангиатных он возникает из одной клетки и имеет однослойную стенку (от греч. leptos – «тонкий»). Обе линии эволюции возникли независимо и обнаружены у древних папоротников. Механизмы вскрывания спорангиев чрезвычайно многообразны. В простейшем случае на вершине спорангия имеется небольшой участок сильно утолщенных клеток – пора. При подсыхании спорангия происходит разрыв. Однако уже у древних папоротников возникло так называемое кольцо – полоска из толстостенных клеток. Кольцо может занимать разное положение: горизонтальное, косое, вертикальное. Примитивным считается сплошное кольцо, или замкнутое, более совершенное – неполное, при котором часть клеток кольца не утолщена (так называемое устье); по нему происходит разрыв стенки (спорангий щитовника мужского – Dryopteris filix-mas) (рис. 4).
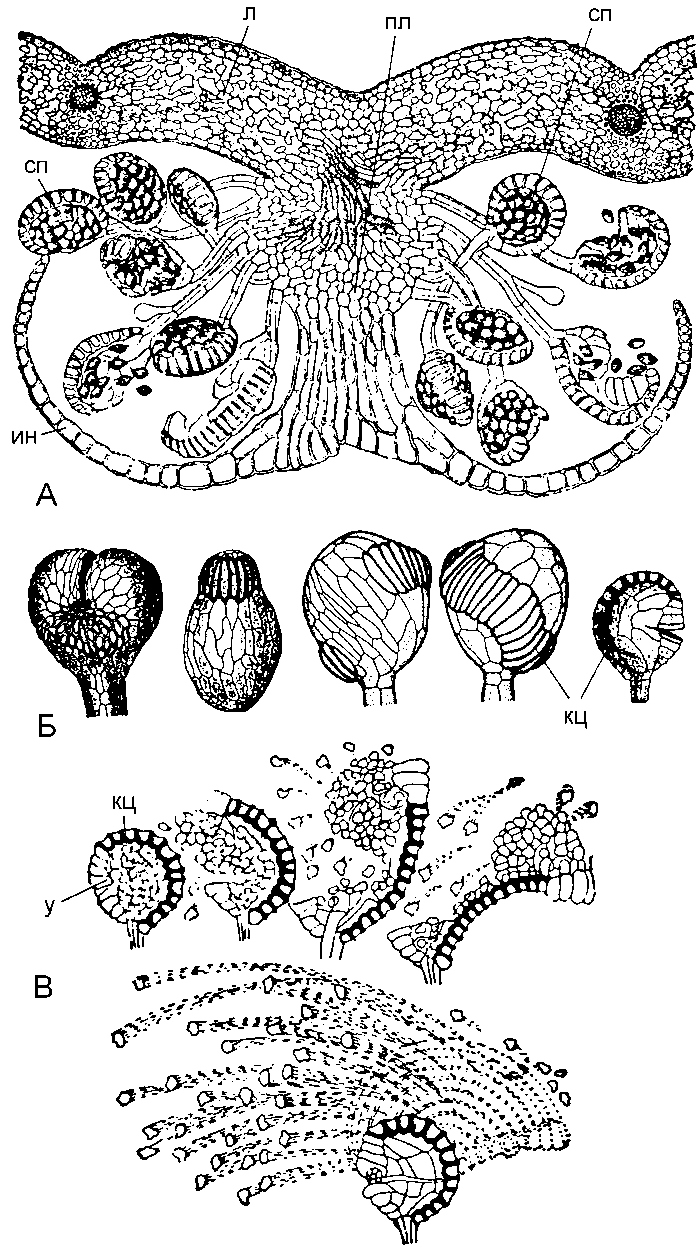
Рис. 4. Строение сорусов и спорангиев папоротников
А – поперечный разрез соруса щитовника мужского (Dryopteris filix - mas); Б – разнообразие спорангиев; В – раскрывание спорангиев; л – лист; пл – плацента; сп – спорангий; кц – кольцо; у – устье; ин – индузий.
У представителей примитивных семейств спорангии крупные, немногочисленные, содержат большое количество спор (8-15 тыс.). В продвинутых семействах – от 64 до 16 спор, в этом случае соблюдается автономность спорангиев и надежность сохранения спор. Через некоторое время (несколько недель или годы) спора прорастает в гаметофит (заросток).
Гаметофиты в основном ведут надземный образ жизни и питаются автотрофно, но у Ужовниковых, Схизейных, Глейхениевых гаметофиты ведут подземное существование, однако могут зеленеть при попадании на поверхность земли. Формирование гаметофита зависит от спектрального состава света: синие лучи способствуют развитию нитчатых, красные – пластинчатых форм.
Рост однослойной пластинки осуществляется за счет краевых клеток, а затем на вершине таллома обособляется инициальная клетка, которая формирует верхушечную меристему, выделяющую специфический гормон – антеридиоген, стимулирующий формирование антеридиев. Встречаются как однополые (мужские, женские), так и смешанные обоеполые заростки. Оплодотворение перекрестное.
Вегетативное размножение широко распространено среди папоротников выводковыми почками на листьях, стеблях, корнях.
В основу классификации папоротников положены разные признаки: анатомическое и морфологическое строение вегетативных органов спорофита, особенности спороношения, характер формирования и строение гаметофита. Относимые ранее к папоротникам (Анейрофитовые и Археоптерисовые) отнесены к Праголосеменным растениям. Отдел Папоротниковидные включает около 300 родов и более 10 тыс. видов.
После мейоза в спорангиях образуются споры, дающие после рассеивания гаметофиты, которые у большинства видов зеленые и независимые в своем питании. Многие из них толщиной всего в один слой клеток, более или менее сердцевидные, с апикальной выемкой; другие толще и могут быть неправильной формы. От нижней поверхности гаметофита отходят, проникая в субстрат, специализированные клеточные нити (ризоиды). На нижней поверхности гаметофита находятся колбовидные архегонии с погруженной в его ткань расширенной нижней частью. Их шейки состоят из нескольких ярусов клеток. Антеридии также находятся на нижней поверхности гаметофита и окружены стерильным покровным слоем.
В них образуются многочисленные, спирально закрученные многожгутиковые сперматозоиды. Когда они созревают, то при достаточном увлажнении антеридии лопаются, высвобождая спермии, заплывающие в шейку архегония. В его брюшке происходит оплодотворение, и возникшая зигота сразу же начинает делиться. Молодой зародыш растет и дифференцируется непосредственно во взрослый спорофит, некоторое время получая питание от гаметофита, но вскоре начиная достаточно интенсивно фотосинтезировать для поддержания своего независимого существования. После укоренения молодого спорофита в почве гаметофит разрушается.
Особняком стоит группа Праголосеменных – Progymnospermae, которая существовала со среднего девона до верхнего карбона.
В группе Праголосеменных прослеживается ряд эволюционных тенденций: 1) увеличение размеров растений, сопровождавшееся усилением деятельности камбия и оживлением древесины за счет паренхимы; 2) переход системы теломов к плосковеткам, а от них – к настоящим листьям; 3) формирование специализированных спороносных органов – спорангиофоров и, как частный случай, спорофиллов; 4) уменьшение количества спорангиев на них до одного; 5) переход от равноспоровости к разноспоровости, сопровождавшийся сокращением количества мегаспор и увеличением их размеров; 6) уменьшение количества мегаспорангиев на мегаспорангиофорах; 7) концентрация вокруг них вегетативных осей, которые способствовали защите мегаспорангиев; 8) формирование на вершине спорангия оттянутого носика, который обеспечивал наиболее эффективное улавливание микроспор.
Вопросы для самостоятельной работы
1. По литературным данным охарактеризуйте ископаемые папоротники: протоптеридиум, кладоксилон, ставроптерис, зигоптерис.
2. Дайте анатомо-морфологическую характеристику класса Мараттиевые (особенности строения спорангиев, сосудов, синангиев).
3. Чем отличаются гаметофиты Мараттиевых от таковых Полиподиопсид?
4. Дайте общую характеристику подклассов Сальвиниевые и Марсилиевые.
5. Объясните, с какими местообитаниями связаны особенности их анатомо-морфологического строения (в т.ч. строение сорусов Сальвиниевых и Марсилиевых)?
6. Расскажите, в чем заключается разноспоровость, редукция гаметофитов Сальвиниевых и Марсилиевых.
Рис. 5. Ужовниковые.
Гроздовник (Botrychium lunaria) – общий вид
У многих Ужовниковых наблюдается способность к эффективному вегетативному размножению. Новые побеги у них появляются из почек, образующихся на корнях. Все Ужовниковые – облигатные (обязательные) микотрофы, но степень зависимости от микоризы различна у разных видов. Гельминтостахис и эпифитные ужовники являются вечнозелеными растениями, но максимум образования новых листьев и период спороношения у них приходятся на определенное время года. У наземных ужовников умеренной зоны листья летнезеленые; в пределах рода Гроздовник есть виды летнезеленые и «зимнезеленые».
Хотя хозяйственного значения Ужовниковые не имеют, эта своеобразная и малочисленная группа заслуживает бережного отношения к себе, а многие виды уже давно нуждаются в охране. Так, вид Botrychium simplex занесен в Красную книгу РФ.
Виды, внесенные в Красную книгу Республики Бурятия (Красная книга Республики Бурятия…, 2002):
Лекция № 5
1. Отдел Папоротниковидные – Polypodiophyta
2. Группа Праголосеменные – Progymnospermae
3. Класс Ужовниковые – Ophioglossopsida
4. Класс Полиподиевые – Polypodiopsida
Отдел Папоротниковидные – Polypodiophyta
Папоротниковидные относят к числу наиболее древних групп высших растений. По своей древности они уступают только Риниофитам и Плауновидным и имеют приблизительно один возраст с Хвощевидными. Однако и по сей день Папоротниковидные продолжают процветать. В настоящее время насчитывается около 300 родов и более 10 тыс. видов Папоротниковидных (рис. 1).

Рис. 1. Папоротникообразные. 1 - Кочедыжник обыкновенный (Athyrium filix - femina Roth.); 2 - Ужовник обыкновенный (Ophioglossum vulgatum L.); 3 - Кочедыжник коричневый (Asplenium trichomanes L.); 4 - Стоножник обыкновенный (Scolopendrium vulgare Sm.); 5 - Дебрянка (Blechnum spicant With.); 6 - Орляк обыкновенный (Pteridium aquilinum Kuhn).
Папоротниковидные распространены очень широко, фактически по всему земному шару, и встречаются в самых различных местообитаниях – начиная с пустынь, кончая болотами, озерами, рисовыми полями и солоноватыми водами.
Но наибольшее разнообразие встречается во влажных тропических лесах. В результате приспособления к столь различным условиям среды у Папоротниковидных выработались очень разные жизненные формы и возникло очень большое разнообразие во внешней форме, внутреннем строении, физиологических особенностях и размерах. Как у большинства высших растений, исключая Моховидные, бесполое поколение – спорофит – является господствующей фазой в их жизненном цикле.
Почти у всех Папоротниковидных спорофит многолетний, однако есть исключение – однолетнее водное или болотное растение Ceratopteris.
Как у большинства высших растений, спорофит Папоротниковидных снабжен корнями, отсутствующими только у части Hymenophyllaceae и у рода Salvinia. Корни придаточные, обычно волокнистые, у Ophioglossaceae – ужовниковых имеют тенденции к мясистости. Ветвление корней моноподиальное.
Стебли никогда не бывают сильно развиты, как правило, листва по массе и размерам преобладает над стеблем. Тем не менее стебли довольно разнообразны по внешнему и внутреннему строению. Прямостоячий стебель древовидных папоротниковидных, несущий на верхушке крону листьев, называют стволом. Высокие стволы обычно снабжены у основания многочисленными воздушными корнями, придающими им устойчивость. Когда стебель ползучий или вьющийся, его называют корневищем, оно может быть длинным или очень коротким, клубневидным, иметь радиальное или дорзовентральное строение. Стебли нередко ветвятся дихотомически или перед листовыми основаниями.
Для классификации имеют значение не только форма и размеры стебля, но и строение тех волосков или чешуй, которыми они покрыты в молодости. Волоски могут быть одноклеточными или однорядными, железистыми (род Cheilanthes), выделять слизь из верхней клетки (род Blechnum). Волоски могут быть ветвистыми, звездчатыми, когда ветви волосков срастаются, образуются чешуи, которые имеют таксономическое значение. Различают чешуи щитовидные (пельтатные) и решетчатые (клатратные). Пельтатные чешуи прикреплены одной из точек своей поверхности, а не краем пластинки. Боковые стенки клеток клатратных чешуй утолщены, образуя явственный решетчатый узор.
Стебли древнейших девонских Папоротниковидных мало отличались от стеблей Риниофитов, и их проводящая система представляла очень примитивную протостелу. У некоторых современных Папоротниковидных сохранилась протостела – Schizaecaea – Схизейных, Гименофилловых и у Gleicheiniaceae– Глейхениевых.
Но у большинства Папоротниковидных проводящая система представлена различными формами сифоностелы. Различают 2 типа: эктофлойную – флоэма окружает ксилему только снаружи, и амфифлойная – флоэма окружает ксилему с обеих сторон, с наружной и с внутренней.
Сифоностела может представлять собой относительно сплошной цилиндр проводящей ткани, но чаще проводящая система представляет сеть проводящих пучков. Промежутки между отдельными пучками заполнены паренхимной тканью. Паренхимные участки, расположенные в стеле над местами отхождения листовых следов, называют листовыми прорывами, или лакунами. Амфифлойную сифоностелу, в которой последовательные листовые прорывы значительно отделены друг от друга, называют соленостелой (от греч. solen – «канал»), если листовые прорывы в амфифлойной сифиностеле расположены столь близко, что нижняя часть одного прорыва параллельна верхней части другого, ее называют диктиостелой (от греч. diktyon – «сеть»). Диктиостела представляет собой сплошной цилиндр, состоящий из сети переплетающихся пучков. Отдельный пучок диктиостелы называют меристемой (от греч. meros – «часть»). Соленостела и диктиостела являются наиболее распространенными типами стелы среди современных папоротников.
Листья папоротников, часто называемые вайями, во многих отношениях отличаются от листьев Хвощевидных и особенно Плауновидных. В то время как листовые органы Плауновидных представляют собой просто выросты на наземных осевых органах, а листья Хвощевидных являются видоизмененными боковыми веточками, у папоротников листья морфологически соответствуют целым крупным ветвям их вероятных предков – Риниофитов. Какие существуют доказательства? Примитивные палеозойские Папоротниковидные имели радиально-симметричные листовые органы, которые лишь с натяжкой можно назвать листьями. Также одиночное и верхушечное расположение спорангиев у наиболее примитивных. Для всех папоротников характерны верхушечный и длительный рост листьев, обычно большие размеры и сложно рассеченная форма листовой пластинки (рис. 2).

Рис. 2. Схема строения листа папоротника
1 – черешок; 2 – пластинка листа; 3 – перо первого порядка; 4 – перышки (или перья второго порядка); 5 – лопасть перышка; 6 – рахис.
Крупные и сложные листовые следы и наличие листовых прорывов в стеле обнаруживаются у большинства видов. Листья папоротников отличаются большим разнообразием. В большинстве случаев листья совмещают две функции – фотосинтез и спороношение. У многих (Matteuccia struthiopteris, Onoclea sensibilis, Drynaria) листья дифференцированы на стерильные и фертильные. Считается, что все 3 типа листьев существовали в раннем палеозое независимо.
У большинства представителей семейства Polypodiaceae черешки соединяются с корневищем посредством особого сочленения или без него (возможность свободно менять ориентацию к источнику света), наличие сочленения является систематическим признаком. Для систематики имеет значение также строение проводящей системы черешка, в частности форма проводящего пучка на поперечном срезе (у осмундовых С-образная форма, у Plagiogyriaceae – Y-образная форма).
Важное значение имеет число проводящих пучков – 1,2 или несколько. У Aspleniaceae обычно 3-7, но у большинства только 2, соединенные наверху в один.
Примитивные листья имеют дихотомическое ветвление (равно- или неравнодихотомическое). У подавляющего большинства листья перистые – однажды, дважды или многократно. Пластинка перистого листа в отличие от дихотомического имеет стержень или рахис (от греч. rhachis – «позвоночник»), представляющий собой продолжение черешка. Стержень соответствует главной жилке целого листа, однорядные сегменты называются перьями (могут быть цельными, лопастными). У папоротников дихотомическое жилкование встречается у примитивных форм – открытое, не образующее сети (представитель семейства Гименофилловых – Trichomanes reniforme). Для более продвинутых форм характерно сетчатое жилкование, которое возникло не сразу.
У Папоротниковидных встречаются почти все основные типы устьичного аппарата (за исключением анизоцитного) – десмоцитный, перицитный, полоцитный. Тип устьичного аппарата специфичен и является систематическим признаком.
Размножение папоротников осуществляется в основном за счет спор. Большинство видов – равноспоровые растения. Для всех папоротников характерно отсутствие специализированных спороносных побегов – стробилов. В основном спорангии сгруппированы в сорусы; у мараттиевых сорусы, срастаясь между собой, образуют синангии. Если у примитивных видов одиночные спорангии располагаются по краям листьев либо на вершинах лопастей, при этом каждый спорангий снабжен самостоятельной жилкой, то у большинства папоротников спорангии или сорусы располагаются на нижней поверхности листьев. Место прикрепления спорангия к листу называется плацентой (ложе). В ходе эволюции происходило разрастание плаценты, и она приобретала продолговатую или шаровидную форму. Кроме этого, спорангии на плаценте располагаются в определенной последовательности, следовательно, созревание и вскрывание неодновременное. Увеличение длины ножки увеличивает количество спорангиев на плаценте. Эти два процесса – разрастание плаценты и увеличение длины ножки спорангия – происходили независимо в разных систематических группах. Остановимся подробнее на строении спорангиев, размещение которых является диагностическим признаком.
Листья папоротников возникли в процессе эволюции из ветвей риниофитовых предков. Очевидно, что спорангии древнейших Папоротниковидных имели верхушечное (терминальное) расположение, т.е. сидели на верхушках конечных веточек дихотомически разветвленного листа. Среди ныне живущих наиболее близкое к верхушечному расположению спорангиев наблюдается у гроздовника (Botrychium), относящегося к примитивным Ужовниковым. Следующий тип расположения спорангиев – краевое (маргинальное), наблюдаемый у многих папоротников, можно рассматривать как результат возникновения листовой пластинки.
Значительным шагом вперед в приспособительной эволюции папоротников был переход на нижнюю сторону листовой пластинки. Наряду с перемещением спорангиев на нижнюю сторону пластинки происходит и другое изменение. Сначала спорангии бывают расположены одиночно (у Схизейных не образуют групп). Но уже у некоторых папоротников с краевым расположением (Dicksonia, Loxsoma) или у всех Гименофилловых спорангии сгруппированы в отдельные спорангиальные кусочки, или сорусы (от греч. soros – «куча»), что дает возможность концентрированного, более эффективного снабжения питательными веществами (рис. 3).
Рис. 3. Расположение спорангиев и сорусов папоротников
1 – спорангии орляка (Pteridium); 2 – сорусы вудсии (Woodsia); 3 – сорусы с покрывальцем щитовника (Dryopteris); 4 – сорусы, встречающиеся у родов Asplenium, Athyrium, Polypodium.
В биологическом отношении чрезвычайно важен порядок развития и созревания спорангиев на листе или в каждом отдельном сорусе. У вымерших Папоротниковидных, и ныне живущих Ужовниковых, Мараттиевых, Осмундовых, Плагиогириевых, Схизейных, Глейхениевых и Матониевых все спорангии развиваются одновременно (внутри соруса), т.е. симультанно (представители семейств Плагиогириевых, Глейхениевых и Матониевых). Сорусы, в которых происходит одновременное развитие и созревание спорангиев, называют простыми. Их существенный недостаток – при ухудшении внешних условий подвергается риску и угнетению произрастание и развитие спор.
Выход – в последовательном развитии и созревании спорангиев в сорусе. Это достигается посредством удлинения плаценты (рецептакула) соруса и увеличения числа спорангиев при уменьшении их размеров.
Устанавливается базипетальная (от греч. basis – «основание», от лат. petere – «устремляться») последовательность, когда развитие начинается с верхней части плаценты (в частности, представители семейства гименофилловых имеют длинное, цилиндрическое ложе). Сорусы с базипетальным развитием называют градатными (от лат. gradatio – «постепенное возвышение, усиление»). Такие сорусы встречаются у Циатейных, Асплениевых, у всех разноспоровых. Смешанные сорусы возникают независимо в разных линиях эволюции.
В дальнейшем произошло важное событие – появились защитные приспособления. Так, в простейших случаях сорусы прикрываются загибающимся краем листа, у более специализированных видов формируется индузий (от лат. indusium – «верхняя туника»). У большинства семейств с простыми сорусами индузий отсутствует.
Другая форма защиты – это срастание в синангии (у некоторых Мараттиевых). Исключение – род Matonia, у которого настоящее покрывало встречается с простыми сорусами.
Индузий имеют папоротники с градатными сорусами. По характеру заложения и строению стенки спорангия папоротники подразделяются на лепто - и эуспорангиатные.
У эуспорангиатных папоротников спорангий возникает из группы клеток и имеет многослойную стенку, у лептоспорангиатных он возникает из одной клетки и имеет однослойную стенку (от греч. leptos – «тонкий»). Обе линии эволюции возникли независимо и обнаружены у древних папоротников. Механизмы вскрывания спорангиев чрезвычайно многообразны. В простейшем случае на вершине спорангия имеется небольшой участок сильно утолщенных клеток – пора. При подсыхании спорангия происходит разрыв. Однако уже у древних папоротников возникло так называемое кольцо – полоска из толстостенных клеток. Кольцо может занимать разное положение: горизонтальное, косое, вертикальное. Примитивным считается сплошное кольцо, или замкнутое, более совершенное – неполное, при котором часть клеток кольца не утолщена (так называемое устье); по нему происходит разрыв стенки (спорангий щитовника мужского – Dryopteris filix-mas) (рис. 4).
Рис. 4. Строение сорусов и спорангиев папоротников
А – поперечный разрез соруса щитовника мужского (Dryopteris filix - mas); Б – разнообразие спорангиев; В – раскрывание спорангиев; л – лист; пл – плацента; сп – спорангий; кц – кольцо; у – устье; ин – индузий.
У представителей примитивных семейств спорангии крупные, немногочисленные, содержат большое количество спор (8-15 тыс.). В продвинутых семействах – от 64 до 16 спор, в этом случае соблюдается автономность спорангиев и надежность сохранения спор. Через некоторое время (несколько недель или годы) спора прорастает в гаметофит (заросток).
Гаметофиты в основном ведут надземный образ жизни и питаются автотрофно, но у Ужовниковых, Схизейных, Глейхениевых гаметофиты ведут подземное существование, однако могут зеленеть при попадании на поверхность земли. Формирование гаметофита зависит от спектрального состава света: синие лучи способствуют развитию нитчатых, красные – пластинчатых форм.
Рост однослойной пластинки осуществляется за счет краевых клеток, а затем на вершине таллома обособляется инициальная клетка, которая формирует верхушечную меристему, выделяющую специфический гормон – антеридиоген, стимулирующий формирование антеридиев. Встречаются как однополые (мужские, женские), так и смешанные обоеполые заростки. Оплодотворение перекрестное.
Вегетативное размножение широко распространено среди папоротников выводковыми почками на листьях, стеблях, корнях.
В основу классификации папоротников положены разные признаки: анатомическое и морфологическое строение вегетативных органов спорофита, особенности спороношения, характер формирования и строение гаметофита. Относимые ранее к папоротникам (Анейрофитовые и Археоптерисовые) отнесены к Праголосеменным растениям. Отдел Папоротниковидные включает около 300 родов и более 10 тыс. видов.
После мейоза в спорангиях образуются споры, дающие после рассеивания гаметофиты, которые у большинства видов зеленые и независимые в своем питании. Многие из них толщиной всего в один слой клеток, более или менее сердцевидные, с апикальной выемкой; другие толще и могут быть неправильной формы. От нижней поверхности гаметофита отходят, проникая в субстрат, специализированные клеточные нити (ризоиды). На нижней поверхности гаметофита находятся колбовидные архегонии с погруженной в его ткань расширенной нижней частью. Их шейки состоят из нескольких ярусов клеток. Антеридии также находятся на нижней поверхности гаметофита и окружены стерильным покровным слоем.
В них образуются многочисленные, спирально закрученные многожгутиковые сперматозоиды. Когда они созревают, то при достаточном увлажнении антеридии лопаются, высвобождая спермии, заплывающие в шейку архегония. В его брюшке происходит оплодотворение, и возникшая зигота сразу же начинает делиться. Молодой зародыш растет и дифференцируется непосредственно во взрослый спорофит, некоторое время получая питание от гаметофита, но вскоре начиная достаточно интенсивно фотосинтезировать для поддержания своего независимого существования. После укоренения молодого спорофита в почве гаметофит разрушается.
Особняком стоит группа Праголосеменных – Progymnospermae, которая существовала со среднего девона до верхнего карбона.






