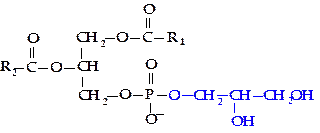Нуклеиновые кислоты представляют собой высокомолекулярные линейные гетерополимеры с молекулярной массой от 2 . 104 до 1,2 . 108 Да, а иногда и более. Мономерными звеньями нуклеиновых кислот являются нуклеотиды — сложные органические молекулы, состоящие из азотистых оснований, остатка пентозы (рибозы или дезоксирибозы) и фосфорной кислоты. В зависимости от типа пентозы нуклеиновые кислоты подразделяются на дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Молекулы нуклеиновых кислот имеют самую различную длину, величина которой составляет от 10 нм до 10-50 мм, причем число нуклеотидов колеблется от 5000 до 5 млн.
Основным местом локализации ДНК являются структуры клеточного ядра — хромосомы, в которых ДНК находится в виде комплексов с белками (гистонами) — хроматина. ДНК (около 1% от общего его количества) также обнаружена в митохондриях всех типов эукариотических клеток и в хлоропластах растительных клеток. В структуре ядерной ДНК заложена информация о видовых специфических признаках, которые определяют характер данной клетки и всего организма и передаются по наследству. В цитоплазме клеток имеются значительные количества РНК, участвующие в реализации генетической информации путем трансляции.
Нуклеиновые кислоты являются многоосновными кислотами, которые при мягком гидролизе щелочами распадаются на мононуклеотиды. При полном гидролизе нуклеиновых кислот образуются азотистые основания, моносахарид пентоза (рибоза или дезоксирибоза) и фосфорная кислота.
Азотистые основания, входящие в состав нуклеиновых кислот, являются производными ароматических гетероциклических соединений — пурина и пиримидина (рисунок 2.18).
Среди пуриновых азотистых оснований в гидролизатах обоих классов нуклеиновых кислот (ДНК и РНК) преимущественно встречаются аденин и гуанин.
Кроме перечисленных пуриновых оснований, в клетках обнаруживают гипоксантин (6-оксопурин) и ксантин (2,6-диоксопурин), которые образуются в результате дезаминирования аденина и гуанина и играют существенную роль в процессах обмена нуклеиновых кислот. Гипоксантин и ксантин в небольших количествах найдены в составе некоторых РНК.
Среди пиримидиновых оснований основное значение имеют цитозин (входит в состав ДНК и РНК), урацил (входит в состав РНК) и тимин (входит в состав ДНК) (рисунок 2.18).
Углеводная часть нуклеотидов, входящих в РНК, представлена рибозой, а входящих в ДНК, — дезоксирибозой. Пентозы в нуклеиновых кислотах всегда присутствуют в b-D-фуранозной форме:

Углеродные атомы пентоз в нуклеотидах нумеруются со знаком «штрих», чтобы их можно было отличить от атомов азотистых оснований.

Рисунок 2.18 - Строение пуриновых и пиримидиновых
Азотистых оснований
Соединения азотистых оснований с пентозой называют нуклеозидами. Нуклеозиды (аденозин, гуанозин, цитидин, тимидин, уридин), выделяемые из нуклеиновых кислот, представляют собой N-гликозиды. Нуклеозиды, содержащие в качестве углеводной части D-рибозу, называют рибонуклеозидами, а содержащие 2-дезокси-D-рибозу — дезоксирибонуклеозидами.
Нуклеотиды — мономерные звенья нуклеиновых кислот — представляют собой монофосфорные эфиры нуклеозидов.
У рибонуклеотидов остаток фосфорной кислоты может находиться в положениях 2', 3' и 5'. В случае дезоксирибонуклеотидов остаток фосфорной кислоты может находиться только в положениях 3' и 5':
Основными фрагментами, полученными из РНК и ДНК, являются следующие мононуклеотиды.
Мононуклеотиды РНК: аденозин-3'- и 5'-фосфаты (адениловые кислоты), гуанозин-3'- и 5'-фосфаты (гуаниловые кислоты), цитидин-3'- и 5'-фосфаты (цитидиловые кислоты), уридин-3'- и 5'-фосфаты (уридиловые кислоты).
Мононуклеотиды ДНК: 2'-дезоксиаденозин-3'- и 5'-фосфаты (дезоксиадениловые кислоты); 2'-дезоксигуанозин-3'- и 5'-фосфаты (дезоксигуаниловые кислоты); 2'-дезоксицитидин- 3'- и 5'-фосфаты (дезоксицитидиловые кислоты); 2'-дезокситимидин-3'- и 5'-фосфаты (тимидиловые кислоты).
Исходя из принятого сокращенного обозначения нуклеозидов (А, Г, Ц, Т, У), монофосфаты принято обозначать АМФ, ГМФ, dAMФ («d» - от дезокси-) и т. д., если фосфат присоединен к углероду 5'- рибозы или дезоксирибозы. Соответствующие монофосфаты с фосфатной группой, присоединенной к третьему атому углерода, обозначаются А-3'-МФ, dA-3'-МФ.
Помимо нуклеотидмонофосфатов, в живых организмах встречаются нуклеотиддифосфаты (например АДФ, УТФ) и нуклеотидтрифосфаты (АТФ, ГТФ), а также циклические нуклеотиды (цАМФ, цГМФ), являющиеся внутриклеточными посредниками различных внеклеточных сигналов (гормонов, нейромедиаторов и т. д.). Особое место занимают нуклеотиды в составе коферментов, являющиеся производными аденозинмонофосфата: никотинамидадениндинуклеотид (НАД+), никотинамидадениннуклеотид-фосфат (НАДФ+), флавинадениндинуклеотид (ФАД+), коэнзим А (КоА) и др.
СТРОЕНИЕ ДНК
Молекулы нуклеиновых кислот всех типов живых организмов — это длинные неразветвленные полимеры мононуклеотидов. Роль мостика между нуклеотидами выполняет 3',5'-фосфодиэфирная связь, соединяющая 5'-фосфат одного нуклеотида и 3'-гидроксильный остаток рибозы (или дезоксирибозы) следующего. В связи с этим полинуклеотидная цепь оказывается полярной. На одном ее конце остается свободной 5'-фосфатная группа, на другом 3'-ОН-группа.
ДНК, подобно белкам, имеет первичную, вторичную и третичную структуры.
Первичная структура ДНК. Данная структура определяет закодированную в ней информацию, представляя собой последовательность чередования дезоксирибонуклеотидов в полинуклеотидной цепи.
Молекула ДНК состоит из двух спиралей, имеющих одну и ту же ось и противоположные направления. Сахарофосфатный остов располагается по периферии двойной спирали, а азотистые основания находятся внутри. Остов содержит ковалентные фосфодиэфирные связи, а обе спирали между основаниями соединены водородными связями и гидрофобными взаимодействиями.
Эти связи впервые были открыты и изучены Э.Чаргаффом в 1945 г. и получили название принципа комплементарности, а особенности образования водородных свзей между основаниями называются правилами Чаргаффа:
пуриновое основание всегда связывается с пиримидиновым: аденин - с тимином (А®Т), гуанин – с цитозином (Г®Ц);
молярное соотношение аденина к тимину и гуанина к цитозину равно 1 (А=Т, или А/Т=1 и Г=Ц, или Г/Ц=1);
сумма остатков А и Г равно сумме остатков Т и Ц, т.е. А+Г=Т+Ц;
в ДНК, выделенных из разных источников, отношение (Г+Ц)/(А+Т), называемое коэффициентом специфичности, неодинаково.
Правила Чаргаффа основаны на том, что аденин образует две связи с тимином, а гуанин образует три связи с цитозином:

На основани правил Чаргаффа можно представить двуспиральную структуру ДНК, которая приведена на рисунке 2.19.
Вторичная структура ДНК. В соответствии с моделью, предложенной в 1953 г. Дж. Уотсоном и Ф. Криком, вторичная структура ДНК представляет собой двухцепочечную правозакрученную спираль из комплементарных друг другу антипараллельных полинуклеотидных цепей.
Для вторичной структуры ДНК решающим являются две особенности строения азотистых оснований нуклеотидов. Первая заключается в наличии групп, способных образовывать водородные связи. Вторая особенность заключается в том, что пары комплементарных оснований А—Т и Г—Ц оказываются одинаковыми не только по размеру, но и по форме.
Благодаря способности нуклеотидов к спариванию, образуется жесткая, хорошо стабилизированная двухцепочечная структура. Основные элементы и параметрические характеристики такой структуры наглядно изображены на рисунке 2.20.
На основе тщательного анализа рентгенограмм выделенных ДНК установлено, что двойная спираль ДНК может существовать в виде нескольких форм (А, В, С, Z и др.). Указанные формы ДНК различаются диаметром и шагом спирали, числом пар оснований в витке, углом наклона плоскости оснований по отношению к оси молекулы (рисунок 2.21).

П р и м е ч а н и е: A-аденин, G-гуанин, C-цитозин, T-тимин
Рисунок 2.19 - Схематическое изображение двуспиральной
молекулы ДНК
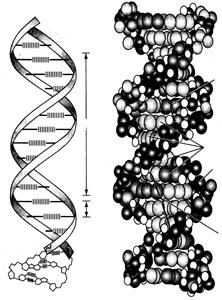
Рисунок 2.20 - Модель двуцепочной спирали ДНК
| 
Рисунок 2.21 - Схематическое
Изображение А- и В-форм
Двойной спирали ДНК
|
Третичная структура ДНК. У всех живых организмов двухспиральные молекулы ДНК плотно упакованы с образованием сложных трехмерных структур.
Двухцепочечные ДНК прокариот, имеющие кольцевую ковалентно-замкнутую форму, образуют левые (—) суперспирали. Третичная структура ДНК эукариотических клеток также образуется путем суперспирализации, но не свободной ДНК, а ее комплексов с белками хромосом (белки-гистоны классов Н1, Н2, Н3, Н4 и Н5) (рисунок 2.22).
 Рисунок 2.22 – Схематическое изображение третичной структуры ДНК
Рисунок 2.22 – Схематическое изображение третичной структуры ДНК
|
В пространственной организации хромосом можно выделить несколько уровней. Первый уровень – нуклеосомный. В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм и укорачивается примерно в 7 раз.
Вторым уровнем пространственной организации хромосом является образование из нуклеосомной нити хроматиновой фибриллы диаметром 20— 30 нм (уменьшение линейных размеров ДНК еще в 6—7 раз).
Третичный уровень организации хромосом обусловлен укладкой хроматиновой фибриллы в петли. В образовании петель принимают участие негистоновые белки. Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 пар нуклеотидов. В результате такой упаковки линейные размеры ДНК уменьшаются примерно в 200 раз. Петлеобразная доменная организация ДНК, называемая интерфазной хромонемой, может подвергаться дальнейшей компактизации, степень которой меняется в зависимости от фазы клеточного цикла.
СТРОЕНИЕ РНК
В цитоплазме клеток содержатся три основных функциональных вида РНК: матричные РНК (мРНК), выполняющие функции матриц белкового синтеза; рибосомные РНК (рРНК), выполняющие роль структурных компонентов рибосом; и транспортные РНК (тРНК), участвующие в трансляции (переводе) информации мРНК в последовательность аминокислот молекулы белка.
В ядре клеток обнаруживают ядерную РНК, составляющую от 4 до 10% от суммарной клеточной РНК. Основная масса ядерной РНК представлена высокомолекулярными предшественниками рибосомных и транспортных РНК. Предшественники высокомолекулярных рРНК (28 S, 18 S и 5 S РНК) в основном локализуются в ядрышке.
РНК является основным генетическим материалом у некоторых вирусов животных и растений (геномные РНК). Для большинства РНК вирусов характерна обратная транскрипция их РНК генома, направляемая обратной транскриптазой.
Все рибонуклеиновые кислоты представляют собой полимеры рибонуклеотидов, соединенных, как в молекуле ДНК, 3',5'-фосфорнодиэфирными связями. В отличие от ДНК, имеющей двухцепочечную структуру, РНК представляет собой одноцепочечные линейные полимерные молекулы.
Строение мРНК. мРНК — наиболее гетерогенный в отношении размеров и стабильности класс РНК. Содержание мРНК в клетках составляет 2—6% от общего количества РНК. мРНК состоят из участков — цистронов, определяющих последовательность аминокислот в кодируемых ими белках.
Строение тРНК. Транспортные РНК выполняют функции посредников (адаптеров) в ходе трансляции мРНК. На их долю приходится примерно 15% суммарной клеточной РНК. Каждой из 20 протеиногенных аминокислот соответствует своя тРНК. Для некоторых аминокислот, кодируемых двумя и более кодонами, существуют несколько тРНК.
тРНК представляют собой сравнительно небольшие одноцепочечные молекулы, состоящие из 70—93 нуклеотидов. Их молекулярная масса составляет (2,4—3,1) . 104 кДа.
Вторичная структура тРНК формируется за счет образования максимального числа водородных связей между внутримолекулярными комплементарными парами азотистых оснований. В результате образования этих связей полинуклеотидная цепь тРНК закручивается с образованием спирализованных ветвей, заканчивающихся петлями из неспаренных нуклеотидов. Пространственное изображение вторичных структур всех тРНК имеет форму клеверного листа (рисунок 2.23).
В «клеверном листе» различают четыре обязательные ветви, более длинные тРНК, кроме того, содержат короткую пятую (дополнительную) ветвь. Адапторную функцию тРНК обеспечивают акцепторная ветвь, к 3'-концу которой присоединяется эфирной связью аминокислотный остаток, и противостоящая акцепторной ветви антикодоновая ветвь, на вершине которой находится петля, содержащая антикодон. Антикодон представляет собой специфический триплет нуклеотидов, который комплементарен в антипараллельном направлении кодону мРНК, кодирующему соответствующую аминокислоту.
Т-Ветвь, несущая петлю псевдоуридина (ТyС-петлю), обеспечивает взаимодействие тРНК с рибосомами.
Д-ветвь, несущая дегидроуридиновую петлю, обеспечивает взаимодействие тРНК с соответствующей аминоацил-тРНК-синтетазой.
 Рисунок 2.23 - Вторичная
структура тРНК
Рисунок 2.23 - Вторичная
структура тРНК
|
 Функции пятой дополнительной ветви пока мало исследованы, вероятнее всего она уравнивает длину разных молекул тРНК.
Функции пятой дополнительной ветви пока мало исследованы, вероятнее всего она уравнивает длину разных молекул тРНК.
  Рисунок 2.24 - Третичная структура тРНК (по А.С. Спирину)
Рисунок 2.24 - Третичная структура тРНК (по А.С. Спирину)
|
Третичная структура тРНК очень компактна и образуется путем сближения отдельных ветвей клеверного листа за счет дополнительных водородных связей с образованием L-образной структуры «локтевого сгиба» (рисунок 2.24). При этом акцепторное плечо, связывающее аминокислоту, оказывается расположенным на одном конце молекулы, а антикодон — на другом.
Строение рРНК и рибосом. Рибосомные РНК формируют основу, с которой связываются специфические белки при образовании рибосом. Рибосомы — это нуклеопротеиновые органеллы, обеспечивающие синтез белка на мРНК. Число рибосом в клетке очень велико: от 104 у прокариот до 106 у эукариот. Локализуются рибосомы главным образом в цитоплазме, у эукариот, кроме того, в ядрышке, в матриксе митохондрий и строме хлоропластов. Рибосомы состоят из двух субчастиц: большой и малой. По размерам и молекулярной массе все изученные рибосомы делят на 3 группы — 70S рибосомы прокариот (S-коэффициент седиментации), состоящие из малой 30S и большой 50S субчастиц; 80S рибосомы эукариот, состоящие из 40S малой и 60S большой субчастиц.
Малая субчастица 80S рибосом образована одной молекулой рРНК (18S) и 33 молекулами различных белков. Большая субчастица образована тремя молекулами рРНК (5S, 5,8S и 28S) и примерно 50 белками.
Вторичная структура рРНК образуется за счет коротких двуспиральных участков молекулы — шпилек (около 2/3 рРНК), 1/3 — представлена однотяжевыми участками, богатыми пуриновыми нуклеотидами.
ЛИПИДЫ, СТРОЕНИЕ И СВОЙСТВА
Липиды (от греч. «lipos» — жир) — низкомолекулярные органические соединения, полностью или почти полностью нерастворимые в воде и хорошо растворимые в неполярных органических растворителях (хлороформ, метанол, эфир, бензол и др.).
Гидрофобность (или липофильность) является общим признаком этого класса соединения. В их состав входят спирты, жирные кислоты, азотистые соединения, фосфорная кислота, углеводы и др.
К основным функциям липидов относятся:
¾ структурная. В комплексе с белками липиды являются структурными компонентами всех биологических мембран клеток. Они влияют на их проницаемость, участвуют в передаче нервного импульса, в создании межклеточного взаимодействия;
¾ энергетическая. Липиды, являясь более восстановленными по отношению к углеводам, служат наиболее энергоемким «клеточным топливом». При окислении 1 г жира выделяется 39 кДж энергии, что в два раза больше, чем при окислении 1 г углеводов;
¾ резервная. Липиды являются наиболее компактной формой депонирования энергии в клетке. Они резервируются в адипоцитах — клетках жировой ткани;
¾ защитная. Обладая выраженными термоизоляционными свойствами, липиды предохраняют организм от термических воздействий; жировая прокладка защищает тело и органы животных от механических и физических повреждений; защитные оболочки в растениях (восковой налет на листьях и плодах) защищают от инфекции и излишней потери или накопления влаги;
¾ регуляторная. Некоторые липиды являются предшественниками витаминов, гормонов, в том числе гормонов местного действия. Кроме того, от состава, свойств, состояния мембранных липидов во многом зависит активность мембрано-связанных ферментов.
Липиды представляют собой разнородные в химическом отношении вещества. В связи с этим существуют разные подходы к их классификации. На рисунке 2.25 приведена классификация липидов, в соответствии с которой они сгруппированы в отдельные классы и группы на основании их химического строения и состава.
Определяющим признаком для первичной классификации липидов, приведенной выше, являются входящие в состав липидов многоатомные алифатические спирты, содержащие две или три гидроксильные группы.
Жирные кислоты. Многообразие и физико-химические свойства липидов в основном обусловлены наличием в их составе жирных кислот.
В природе обнаружено более 200 жирных кислот. Однако широкое распространение имеют не более 20, которым присущ ряд общих свойств и особенностей:

Рисунок 2.25 - Классификация липидов
¾ жирные кислоты, входящие в состав липидов высших растений и животных — это монокарбоновые кислоты, содержащие линейные углеводородные цепи (обычно С12—С20) с общей формулой СН3(СН2)nСООН;
¾ жирные кислоты обычно содержат четное число атомов углерода (n – кратно 2). Однако в природе встречаются также кислоты с нечетным числом углеродных атомов;
¾ жирные кислоты могут быть как насыщенными, т.е. содержат в углеводородной цепочке только ковалентные связи, так и ненасыщенными, т.е. содержат одну и более ненасыщенных (этиленовых) связей. Они всегда разделены одной метиленовой группой:
—СН=СН—СН2—СН=СН—
Необходимо отметить, что на долю ненасыщенных кислот в природных липидах приходится примерно 3/4 всех жирных кислот.
¾ природные ненасыщенные жирные кислоты чаще имеют цис -конфигурацию, крайне редко в полиеновых кислотах встречается транс -конфигурация:

В таблице 2.4 приведены названия и структурные формулы некоторых наиболее распространенных высших жирных ксилот.
Таблица 2.4 - Некоторые физиологически важные высшие жирные кислоты
| Число атомов
углерода в цепи
| Тривиальное название
| Систематическое название
|
| Насыщенные жирные кислоты
|
| С16
| пальмитиновая
| гексадекановая
|
| СН3-(СН2)14-СООН
|
| С18
| стеариновая
| окстадекановая
|
| СН3-(СН2)16-СООН
|
| Моноеновые жирные кислоты
|
| С18
| олеиновая
| 9-октадеценовая
|
| СН3-(СН2)7-СН=СН-(СН2)7-СООН
|
| Полиеновые жирные кислоты
|
| С18
| линолевая
| 9,12-октадекадиеновая
|
| СН3-(СН2)4-СН=СН- СН2-СН=СН- (СН2)7-СООН
|
| С18
| линоленовая
| 9,12,15-октадекатриеновая
|
| СН3-СН2-СН=СН- СН2-СН=СН- СН2-СН=СН- (СН2)7-СООН
|
| С20
| арахидоновая
| 5,8,11,14-эйкозатетраеновая
|
| СН3-(СН2)4-СН=СН- СН2-СН=СН- СН2-СН=СН- СН2-СН=СН- (СН2)3-СООН
|
Большое число неполярных связей С—С и С—Н в углеводородной цепи жирных кислот придает неполярный характер молекуле липида в целом, хотя в ней имеется полярная, заряженная, группа — СОО—. Неполярность высших жирных кислот является причиной нерастворимости липидов в воде.
Цис -конфигурация двойной связи придает углеводородной цепи укороченный вид за счет ее изгиба. Введение цис -этиленовой связи существенно влияет на свойства жирных кислот. Так, например, с увеличением числа двойных связей значительно снижается температура плавления жирных кислот, возрастает их растворимость в неполярных растворителях.
Линолевая, линоленовая и другие полиеновые кислоты не синтезируются в организме высших животных и человека и должны поступать в организм с пищей. В связи с тем, что эти кислоты необходимы для нормальной жизнедеятельности организма, их относят к незаменимым (эссенциальным) жирным кислотам или чаще комплекс этих кислот объединяют в группу витаминов F.
Особая роль в организме принадлежит 20-углеродным (эйкозановым) ненасыщенным кислотам (арахидоновой и дигомо-g-линоленовой), являющимся предшественниками тканевых гормонов (эйкозаноидов, простагландинов, тромбоксанов и лейкотриенов).
Ацилглицеролы, или нейтральные липиды, — наиболее распространенная в природе группа липидов. Эти соединения представляют собой сложные эфиры жирных кислот и трехатомного спирта глицерола (глицериды), в котором могут быть этерифицированы одна, две или три гидроксильные группы глицерола с образованием соответственно моно-, ди- и триацилглицеролов:

В природе наиболее часто встречаются триацилглицеролы. Поскольку все приведенные выше ацилглицеролы не содержат ионных групп, они относятся к нейтральным липидам. Если все три кислотных радикала принадлежат одной и той же жирной кислоте, то такие триацилглицеролы называют простыми, если же разным жирным кислотам — то смешанными.
Животные жиры обычно содержат значительное количество насыщенных жирных кислот, благодаря чему они при комнатной температуре остаются твердыми. Жиры, в состав которых входит много ненасыщенных кислот, будут при этих условиях жидкими - их называют маслами.
Для характеристики свойств жира используют константы, или жировые числа, — кислотное число, число омыления, йодное число.
Нейтральные липиды играют важную роль в процессах метаболизма в организме. Триацилглицеролы жировой ткани являются самой компактной и энергоемкой формой хранения энергии, кроме того выполняют в подкожном слое роль физической защиты, термо- и электроизоляторов.
Воска — сложные эфиры высших жирных кислот и высших моноатомных или двухатомных спиртов.
Помимо эфиров, воска содержат свободные высшие жирные спирты, например цетиловый спирт и другие спирты с четным числом углеродных атомов (от С22—С32), а также свободные жирные кислоты с длинной углеводородной цепью (от С14 до С34).
Спермацет, получаемый из головного мозга кашалотов, является сложным эфиром цетилового спирта (первичный спирт, соответствующий пальмитиновой кислоте) и пальмитиновой кислоты.
Воска выполняют в организме преимущественно защитную функцию, которая сводится к образованию защитных покрытий. Они входят в состав жира, покрывающего кожу, шерсть, перья. У растений около 80% от всех липидов, образующих пленку на поверхности листьев, составляют воска. Известно также, что воска являются нормальными метаболитами некоторых микроорганизмов.
Фосфолипиды. Общий признак всех фосфолипидов — наличие в их составе фосфорной кислоты. В зависимости от спиртового компонента они делятся на глицерофосфолипиды и сфингофосфолипиды.
Глицерофосфолипиды.
Общим структурным фрагментом всех глицерофосфолипидов является фосфатидная кислота (1,2-диацил,3-фосфоглицерол):

Молекулы фосфолипидов имеют гидрофобную часть, образованную радикалами жирных кислот, и гидрофильную — остатки фосфорной кислоты, аминокислот, аминоспиртов.
Фосфатидилхолин (лецитин). В своем составе содержит аминоспирт холин (гидроксид 3-гидроксиэтилтриметиламмония). В зависимости от того, с каким атомом углерода глицерола связана фосфорная кислота (a— в крайнем положении, b — в срединном), различают два типа фосфатидилхолинов (a и b).
Фосфатидилхолины (ФХ) широко распространены в клетках, особенно мозговой ткани человека и животных; в растениях они встречаются в соевых бобах, семенах подсолнечника, зародышах пшеницы.
Фосфатидилэтаноламин (кефалин). В состав фосфатидилэтаноламинов (ФЭА) вместо холина входит азотистое основание этаноламин.
ФХ и ФЭА являются главными липидными компонентами мембран клеток.
В результате гидролиза при действии специфичного фермента — фосфолипазы А2 в ФХ или ФЭА возможно отщепление остатка жирной кислоты с образованием лизоформ: лизофосфатидилхолина (ЛФХ) и лизофосфатидилэтаноламина (ЛФЭА):
Фосфатидилсерин. В молекуле фосфатидилсерина полярной группой является остаток аминокислоты серина.
Фосфатидилсерины (ФС) распространены менее широко, чем ФХ и ФЭА. Значение ФС определяется тем, что он является предшественником в синтезе двух других групп.
Плазмалогены. Известны также глицерофосфолипиды, которые в отличие от приведенных выше — ФХ, ФЭА, ФС — отличаются тем, что вместо остатка кислоты при атоме углерода С, они содержат a-, b-ненасыщенный спирт, образующий простую эфирную связь с гидроксильной группой глицерола.
При гидролизе этой эфирной связи образуется альдегид соответствующего спирта. Отсюда название группы — фосфатидали: фосфатидальэтаноламин, фосфатидальхолин, фосфатидальсерин.
 фосфатидальэтаноламин (плазмалоген)
фосфатидальэтаноламин (плазмалоген)
|
На долю плазмалогенов приходится до 10% фосфолипидов мозга и мышечной ткани. Они обнаружены также в эритроцитах, тканях некоторых беспозвоночных (до 25%), входят в состав бактериальных мембран, но практически не встречаются в растениях.
Фосфатидилинозитол. В отличие от других групп глицерофосфолипидов в состав фосфатидилинозитола вместо азотсодержащих соединений входит шестиуглеродный циклический спирт инозитол, представленный одним из его стереоизомеров — моноинозитолом.
Фосфатидилинозитол входит в состав клеточных мембран животных, высших растений, различных микроорганизмов, особенно высоко его содержание в миелиновых оболочках нервных волокон.
Важное значение имеют фосфорилированные производные фосфатидилинозитолов.
Фосфатидилглицерол. Так же как фосфатидилинозитол, фосфотидилглицерол не содержит азотсодержащего соединения. В этих фосфолипидах полярной группой служит еще одна молекула глицерола.
Фосфатидилглицеролы и их аминокислотные производные в значительном количестве содержатся в бактериальных мембранах, а также хлоропластах растений.
Кардиолипин. Этот фосфолипид можно рассматривать как производное фосфатидилглицерола, в котором 3-гидроксигруппа второго остатка молекулы глицерола этерефицирована молекулой фосфатидной кислоты.

Кардиолипин локализован почти исключительно в митохондриях и играет важную роль в структурной организации и функционировании дыхательных комплексов. Кардиолипин является также обязательным компонентом бактериальных клеточных мембран и хлоропластов растений.
Сфингофосфолипиды.
В составе соединений этого класса глицерола нет. Они являются производными сложного ненасыщенного дигидроксиаминоспирта — сфингозина или его насыщенного аналога — дигидросфингозина.
При ацилировании Н2N-группы сфингозина жирной кислотой образуется соединение — церамид, фосфохолиновое производное которого называется сфингомиелином. Сфингомиелины являются наиболее распространенными сфинголипидами, находятся в мембранах животных и растительных клеток. Особенно богата ими нервная ткань. Сфингомиелины обнаружены также в ткани почек, печени и других органов, входят в состав липидов крови.
Гликолипиды (гликосфинголипиды). Липиды этого класса, подобно сфингомиелинам, являются производными церамидов, спиртовая группа которых гликозилирована остатками одного или нескольких углеводов. В зависимости от состава углеводного компонента их делят на цереброзиды и ганглиозиды.
Цереброзиды – это церамидомоносахариды, к ним относятся галактозил-церамиды и глюкозилцерамиды. Гликозидная связь с углеводом имеет обычно Р-конфигурацию.
Галактозилцерамид является основным гликолипидом мозговой и нервной тканей, содержит различные жирные кислоты, в том числе цереброновую (С24), являющуюся гидроксикислотой. В результате сульфатирования галактозилцерамида он превращается в сульфатид.
Ганглиозиды представляют собой более сложные сфинголипиды, чем цереброзиды. В их состав входят сфингозин, жирная кислота, один или несколько углеводов (D-глюкоза, D-галактоза, N-ацетилглюкозамин, N-ацетилгалактозамин, N-ацетилнейраминовая кислота и др.).
Ганглиозиды в больших количествах находятся в нервной ткани. В сером веществе мозга ганглиозиды составляют около 6% мембранных липидов. Ганглиозиды выполняют рецепторные функции. Они активно участвуют в контроле и регуляции межклеточных контактов, рецепции.
Стероиды являются производными циклопентанпергидрофенантрена, содержащего три нелинейно конденсированных насыщенных циклогексановых и одно циклопентановое кольцо.
К стероидам относится большое количество биологически важных соединений: собственно стеролы (или стерины), витамины группы D, поло-
 циклопентанпергидрофенантрен
циклопентанпергидрофенантрен
|
вые гормоны, гормоны коры надпочечников, зоо- и фитоэкдистероидные гормоны, сердечные гликозиды, растительные сапонины и алкалоиды, некоторые яды и др.
Классификация стероидов основана на структуре заместителя у С-17 углеродного атома (длина углеродной цепи), числе и положении гидроксигрупп, степени ненасыщенности.
Наиболее известный среди стеролов — холестерол, содержащийся почти во всех тканях организма. Особенно много его в центральной и периферической нервной системе, подкожном жире, почках и др. Холестерол является одним из главных компонентов цитоплазматической мембраны, а также липопротеинов плазмы крови. В липопротеиновых фракциях крови примерно только одна треть его находится в виде спирта, а две трети — в форме эфиров жирных кислот (холестеридов):
| холестерид (эфир холестерола)
| | 
|
Холестерол служит исходным предшественником для синтеза всех стероидов, функционирующих в организме, — половых гормонов и гормонов коркового слоя надпочечников (кортикостероидов), желчных кислот, витамина D3. Кроме этого, холестерол, входящий в состав клеточных мембран, оказывает существенное в