ОГЛАВЛЕНИЕ
Перечень условных обозначений …………………………………………….... 4
| ВВЕДЕНИЕ ……………………………………………………………………..
|
|
| ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ…………………………………………..
|
|
| РАЗДЕЛ 1. ОБЩАЯ ХАРАКТЕРИСТИКА ВИРУСА ГЕПАТИТА С….
|
|
| 1.1.1 Морфо-биологическая характеристика вируса гепатита С ………….
|
|
| 1.1.2 Структурная организация генома вируса гепатита С и кодированные им белки ………………………………………………………...
|
|
| 1.1.3 Классификация и распространение вируса гепатита С ………………
|
|
| 1.1.4 Морфогенез вирусных частиц гепатита С …………………………….
|
|
| 1.1.5 Основы генетической изменчивости вирусного генома гепатита С...
|
|
| РАЗДЕЛ 2. ДИАГНОСТИКА ГЕПАТИТА С ……………………………....
|
|
| 1.2.1 Основные пути инфицирования вирусом гепатита С ………………..
|
|
| 1.2.2 Биохимические маркеры и симптомы гепатита С ……………………
|
|
| 1.2.3 Методы лабораторной диагностики гепатита С ……………………...
|
|
| 1.2.3.1 Иммунологические методы исследования вируса гепатита….
|
|
| 1.2.3.2 Генетические методы исследования вируса гепатита С ……..
|
|
| 1.2.4 Значение определения генотипического разнообразия изолятов вируса гепатита С ………………………………………………………………..
|
|
| ВЫВОДЫ ……………………………………………………………………...
|
|
| ГЛАВА 2. МЕТОДЫ ИССЛЕДОВАНИЙ…………………………………..
|
|
| 2.1 Забор материала …………………………………………………………..
|
|
| 2.2 Выделение РНК вируса гепатита С ……………………………………...
|
|
| 2.3 Выявление РНК вируса гепатита С ПЦР «в режиме реального времени» …………………………………………………………………………
|
|
| ГЛАВА 3. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ ………
|
|
| 3.1 Результаты исследования сывороток крови на наличие ВГС методом ПЦР «в режиме реального времени» …………………………………………..
|
|
| 3.2 Результаты исследований сывороток крови на наличие ВГС методом ИФА ………………………………………………………………………………
|
|
| 3.3 Сравнительный анализ методов ИФА и ПЦР «в режиме реального времени» при выявлении вируса гепатита С …………………………………..
|
|
| ЗАКЛЮЧЕНИЕ ………………………………………………………………...
|
|
| БИБЛИОГРАФИЧЕСКИЙ СПИСОК ……………………………………....
|
|
| ПРИЛОЖЕНИЕ А……………………………………………………………..
|
|
ПЕРЕЧЕНЬ СОКРАЩЕНИЙ И УСЛОВНЫХ ОБОЗНАЧЕНИЙ
IgG – иммуноглобулин класса G;
IgМ – иммуноглобулин класса М;
ВГС – вирус гепатита С;
ВКО – внутренний контроль образца;
ВОЗ – Всемирная организация здравоохранения;
ГС – гепатит С;
ИФА – иммунно-ферментный анализ;
ОКО – отрицательный контроль образца;
ПКО – положительный контроль образца;
ПЦР – полимеразная цепная реакция;
СПКО – слабоположительный контроль образца;
ВВЕДЕНИЕ
Актуальность проблемы гепатита С обусловлена весомым социально-экономическим ущербом и эпидемической значимостью этого заболевания, повсеместным распространением, тяжестью течения, высокой частотой неблагоприятных исходов инфекции, активным вовлечением в эпидемический процесс лиц репродуктивного и трудоспособного возраста. По оценкам экспертов Европейской ассоциации по изучению болезней печени(Париж, 2005) более 500 млн человек в мире инфицированы гепатитом С. Только хронической формой инфекции страдают 150-170 млн жителей планеты [1, 2].
В настоящее время не вызывает сомнения тот факт, что именно хронические и латентные варианты гепатита С определяют основную часть эпидемического процесса, социальную значимость и прогноз данной инфекции [1].
Учитывая чрезвычайную сложность структуры эпидемического процесса и многофакторность его развития, закономерности течения этой инфекции в полной мере не раскрыты до настоящего времени. На течение инфекционного процесса гепатита С в постоянно изменяющихся социальных и природных условиях большое влияние оказывает значительная генетическая гетерогенность вируса [3, 4, 5, 6, 7, 8, 9, 10, 11, 12].
Сравнение молекулярной вариабельности изолятов, выявляемых в разных популяционных группах на конкретной территории, может использоваться для нужд эпидемиологического надзора и в клинической практике. Несмотря на международные усилия по созданию серологических тест-систем для выявления маркеров инфекции, в настоящее время нет равно чувствительных иммунологических тестов к различным генотипам вируса гепатита С. Такая ситуация диктует необходимость использования молекулярно-генетических методов для изучения циркуляции различных генетических вариантов вируса гепатита С, установления источников инфекции, изучения путей передачи, прогнозирования эволюционных изменений эпидемиологической ситуации для предотвращения дальнейшего распространения гепатита С.(диссертация)
Целью дипломной работы является изучение морфо-биологической и генетической характеристики вирусного гепатита С и методов его диагностики.
Для выполнения поставленной цели необходимо решить следующие задачи:
1. изучить морфологическую и генетическую характеристику вируса гепатита С на основе литературных данных;
2. изучить клиническую картину гепатита С;
3. освоить методы диагностики вируса гепатита С;
4. провести сравнительный анализ ИФА и ПЦР «в режиме реального времени» при выявлении вирусного гепатита С.
ГЛАВА 1.
ОБЗОР ЛИТЕРАТУРЫ
Рисунок 1.3 – Строение вириона вируса гепатита С
Одна открытая рамка считывания кодирует полипротеин длиной 3008-3037 а.о., который процессируется комбинацией вирусных протеиназ и протеиназ клетки-хозяина. На 5'- и 3'-концах генома вируса находится не транслируемый регион (5'- и 3'-NTR) [12, 14, 15].
Протяженность 5'-NTR РНК ВГС составляет около 340 нуклеотидов. Функция этого участка заключается в инициации трансляции. Специфически связываясь с рибосомами (благодаря наличию внутреннего рибосомального сайта связывания) и факторами трансляции клетки-хозяина, он направляет рибосому к инициирующему кодону (AUG) в позиции 342, после чего начинается синтез полипротеина [15, 21].
В 3'-NTR-части РНК вируса гепатита С выделяют три отдела: первый – протяженностью в 28-42 нуклеотида, второй – содержащий поли-U-/поли-А-тракт, и третий – терминальный, высококонсервативный участок протяженностью в 98 нуклеотидов [15]. По аналогии с другими флавивирусами можно предположить, что этот участок играет определенную роль в репликации вируса и упаковке нуклеиновой кислоты в вирусную частицу. Наличие в 3'-NTR-регионе РНК вируса гепатита С различных изолятов высококонсервативного участка нуклеиновых кислот определяет интерес к этому участку в связи с перспективой разработки эффективных лекарственных препаратов [14, 21].
Участок РНК вируса гепатита С, примыкающий к 3'-концу 5'-NTR- региона, протяженностью в 573 нуклеотида, кодирует белок, формирующий капсид вируса (С, или Core) (рис. 1.3). Его С-терминальный регион является необходимым для правильной укладки полипептидной цепи [22].
Выявлено три формы С-белка. Полноразмерная (р21) с молекулярной массой 21 кДа и усеченная (р19), которые обнаружены в мембране эндоплазматического ретикулума. Форма (р16) С-белка, обнаруженная в ядрышках инфицированных гепатоцитов [14, 21]. Он может оказывать влияние на человеческие онкогены, задействованные в развитии гепатоклеточной карциномы у пациентов с хроническим гепатитом С путем супрессии отдельных генов клетки-хозяина. Считается, что С-белки р16 и р21 разнонаправленно влияют на транскрипционную активность опухолевого репрессора р53: первый блокирует, а второй усиливает ее. Таким образом, специфически подавляется апоптоз инфицированных клеток [15]. Кроме того, капсидный белок играет основную роль в индукции митохондриальной дисфункции [23].
На своей поверхности С-белок несет различные высоко-консервативные В-клеточные эпитопы, существование которых чрезвычайно важно для выявления анти-ВГС в процессе лабораторной диагностики инфекции. Core-белок обладает способностью связываться с вирусной РНК, а также выполняет регуляторную роль в репликации вируса [14, 21, 24].
В Е1- и Е2-регионах РНК вируса гепатита С регистрируется значительная генетическая изменчивость и содержится информация о белках оболочки вируса, участвующих в проникновении вируса в клетку, в развитии иммунитета и ускользании от иммунного ответа организма за счет отбора мутантов, способных уклоняться от действия нейтрализующих антител [14, 15]. Кроме того, на мышиной модели показано, что участок Е1-Е2 совместно с Сore ускоряет опухолеобразование, блокируя способность клеток к апоптозу [25].
Зоны генома Е1 и Е2 кодируют белки с молекулярной массой 31 и 70 кДа соответственно. Имеются две формы белка Е2: gp70A (E2) и gp70B (E2-p7). Последний в структуре вируса не обнаружен, предполагается его участие в высвобождении вновь синтезированного вириона вируса из инфицированной клетки [14, 15]. Белки Е1 и Е2 сильно гликозилированы и содержат С-терминальный гидрофобный домен, функционирующий как мембранный якорь. Их созревание происходит при удалении С-концевой последовательности сигнальными пептидазами клетки [24]. Установлено, что при отсутствии белка Е2 происходит нарушение пространственной структуры оболочечного белка Е1 [14].
В неструктурной зоне, расположенной ближе к 3'-концу РНК вируса гепатита С, выделяют участки NS2, NS3, NS4A, NS4B, NS5A и NS5B. В отличие от других флавивирусов, в РНК вируса гепатита С отсутствует первая зона, кодирующая неструктурные белки (NS1), а к Е2-региону примыкает зона NS2 (рис. 1.4). Большинство белков, кодированных неструктурными зонами РНК вируса гепатита С, необходимо для репликации вируса [14, 15]. Антитела, вырабатываемые на неструктурные белки, не обладают полноценными протективными свойствами в отношении вируса гепатита С вследствие его высокой генетической вариабельности [24, 26, 27]. NS2 протеин является трансмембранным белком с молекулярной массой 23 кДа [14]. Его С-конец смотрит в просвет цистерн эндоплазматического ретикулума, N-конец – в цитозоль. Этот белок является цинк-зависимой аутопротеазой, разрезающей NS2 и NS3 белки [15].

Рисунок 1.4 – Организация генома вируса гепатита С
NS3 протеин, с молекулярной массой 70 кДа, выполняет несколько различных функций: является сериновой протеазой (амино-терминальная часть), отщепляя от полипротеина все остальные неструктурные белки (NS3/NS4A, NS4A/NS4B, NS4B/NS5A и NS5A/NS5B); обладает хеликазной и нуклеотидтрифосфатазной активностью [14, 15].
При репликации вируса NS3 белок связывается с поли-U последовательностью на 3'-конце вирусного генома своим РНК-связывающим доменом и затем происходит раскручивание двунитевой РНК. Одновременно идет гидролиз дезоксирубонуклеотидтрифосфатов, осуществляемый другим доменом NS3 [15].
P. Brunetal. (2010) в своей работе показал, что NS3 протеаза может ингибировать сигнальные пути трансдукции врожденного иммунитета. Белок способен специфически взаимодействовать с каталитической субъединицей клеточной протеинкиназы А, участвующей в передаче клеточных сигналов, влияя на переход клетки в состояние неконтролируемого роста. C другой стороны, взаимодействие NS3 с иммунной системой может влиять на течение заболевания либо со спонтанным очищением от вируса, либо развитием хронической инфекции [15, 28].
В NS4 регионе выделяют две зоны – NS4A и NS4B, которые кодируют два гидрофобных белка с молекулярными массами 8 и 26 кДа [14, 15]. NS4A протеин является кофактором для NS3 протеазы, образуя с NS3 белком единый комплекс и выполняя функцию якоря, удерживающего на мембране ядра клетки репликативный комплекс вируса гепатита С [14]. Он также необходим для гиперфосфорилирования NS5A протеина. Функция NS4B протеина остается неясной, предполагается, что он также принимает участие в формировании репликативного комплекса [15]. NS5 регион полипротеина построен из двух больших белков — NS5A (56 кДа) и NS5B (65 кДа). Они освобождаются из полипротеина с помощью NS3-NS4A протеазного комплекса.
NS5A белок высококонсервативен, на ядерной мембране инфицированных клеток он совместно с NS5B образует мембранно-связанный репликативный комплекс. NS5A протеин участвует в процессах фосфорилирования и играет важную роль в формировании механизмов устойчивости клеток к действию интерферона. Предположительно белок NS5A ингибирует один или несколько клеточных белков (например, взаимодействует с интерферон-индуцированной клеточной протеинкиназой), участвующих в начальных стадиях противовирусного действия интерферона [12, 14].
NS5B протеин консервативен и является РНК-зависимой РНК-полимеразой с молекулярной массой 68 кДа, обеспечивая репликацию/ транскрипцию генома вируса [15]. Изучение функциональной роли вирусных белков, кодированных неструктурной зоной РНК вируса гепатита С, прежде всего, важно для поиска новых лечебных препаратов, которые могли бы эффективно блокировать репликативную активность вируса.
Выводы по главе 1
Гепатит С – это инфекционное заболевание, вызываемое РНК-содержащим гепатотропным вирусом.
Гепатит С – инфекция, которая характеризуется первичной гепатотропностью, полиморфизмом клинических проявлений (феномен «айсберга»), длительным бессимптомным течением, исключительно высокой частотой хронизации (до 75-83%) с возможным переходом в цирроз (у 26-35% лиц с хроническим гепатитом С) и первичный рак печени (у 30-40% больных циррозом).
 Вирус гепатита С представляет собой небольшой (55-65 нм в размере) сферический РНК-содержащий вирус из семейства Flaviviridae. ВГС является причиной развития гепатита С в организме человека.
Вирус гепатита С представляет собой небольшой (55-65 нм в размере) сферический РНК-содержащий вирус из семейства Flaviviridae. ВГС является причиной развития гепатита С в организме человека.
ВГС классифицируют по следующим генотипам (основным типам и подтипам): 1 (1a, 1b, 1 c), 2 (2a, 2b, 2c), 3 (3a, 3b), 4 (4a, 4b, 4c, 4d, 4e), 5
(5a), 6 (6a), 7 (7a, 7b), 8 (8a, 8b), 9 (9a), 10 (10a), 11 (11a).
Для заражения гепатита С, по сравнению с гепатитом В, необходима большая инфицирующая доза, поскольку концентрация вирусных частиц при гепатите С не превышает 4lg на 1 мл крови.
Для определения РНК ВГС применяют 5 молекулярно-генетических методов исследования, один из которых является ПЦР «в режиме реального времени».
Рисунок 2.1 – Встраивание интеркалирующих красителей в двухцепочечную ДНК
2. С использованием ДНК-зондов. В реакционную смесь добавляют ДНК зонды, в состав которых входит флуоресцентная метка и гаситель флуоресценции. Когда зонд находится в растворе, гаситель поглощает излучение и свечение отсутствует. В ходе ПЦР происходит присоединение ДНК-зонда к комплиментарной цепи ДНК, одновременно разъединяются флуоресцентная метка и гаситель, что приводит к увеличению детекции свечения (рисунок 2.2).

Рисунок 2.2 – Встраивание ДНК зонда в строящуюся цепь
Интенсивность сигнала пропорциональна концентрации конечного продукта ПЦР. Важнейшей особенностью метода является синхронизация регистрации и амплификации. Это дает возможность оценить кинетику процесса, которая зависит от начального количества исследуемого наследственного материала. Детекция флуоресцентного сигнала от каждого красителя происходит в определенном для него диапазоне длин волн (канале). Диапазон выбирается таким образом, чтобы детектировать сигнал только одного красителя, избегая перекрывания соседних сигналов. Если настроить прибор для детекции сигнала в максимуме флуоресценции каждого красителя, тогда остальные красители в этом максимуме будут иметь очень низкую флуоресценцию, которой можно пренебречь (рисунок 2.3).
Флуоресцентные красители обеспечивают флуоресценцию, прямо пропорциональную количеству ПЦР-продукта репортерную флуоресценцию. Механизмы генерации репортерной флуоресценции различаются в зависимости от типа ПЦР «в реальномвремени».

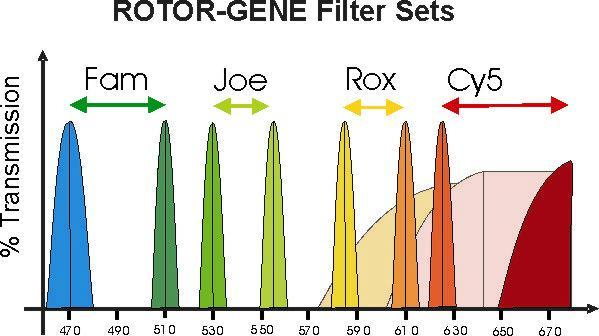
Рисунок 2.3 – Спектр излучаемой флуоресценции различных красителей
На рисунке 2.4 показан типичный образец кинетической кривой ПЦР «в реальном времени». График имеет сигмовидную форму и демонстрирует 3 стадии реакции:
1) инициация (отсутствие детектирующего сигнала, выявляющего ПЦР продукты);
2) экспоненциальная (экспоненциальная зависимость интенсивности флуоресценции от количества циклов ПЦР);
3) насыщения (пороговый уровень флуоресценции).

Рисунок 2.4 – Кинетическая кривая ПЦР «в реальномвремени»
В начальных циклах ПЦР существуют небольшие изменения флуоресцентного сигнала, значения которого определяют исходный (базовый) уровень графика. Увеличение уровня флуоресценции сверх базового свидетельствует о накоплении ПЦР продуктов. При этом фиксируемое пороговое значение флуоресценции может быть установлено выше ее базового уровня. Пороговый цикл определяют как номер цикла, при котором флуоресценция преодолевает фиксированный порог.
Теоретически в конце каждого цикла должно происходить удвоение числа продуктов ПЦР. Результатом циклического процесса является экспоненциальное увеличение количества специфического фрагмента ДНК, которое можно описать формулой (2.1):

| (2.1)
|
где  – количество специфических (ограниченных праймерами) продуктов реакции амплификации;
– количество специфических (ограниченных праймерами) продуктов реакции амплификации;
 – начальное количество ДНК-мишеней;
– начальное количество ДНК-мишеней;
n – число циклов амплификации.
Реальное значение эффективности отдельных циклов амплификации составляет по некоторым данным 78-97%. В случае присутствия в пробе ингибиторов реакции это значение может быть намного меньше, поэтому фактическое количество специфических продуктов амплификации лучше описывает формула (2.2):

| (2.2)
|
где  – значение эффективности реакции.
– значение эффективности реакции.
Следует заметить, что в процессе амплификации на исходной цепи синтезируются и длинные фрагменты, однако их накопление происходит лишь в арифметической прогрессии по формуле (2.3):

| (2.3)
|
где  - количество длинных продуктов амплификации.
- количество длинных продуктов амплификации.
Таким образом, специфические фрагменты, ограниченные на концах праймерами, впервые появляются в конце второго цикла, накапливаются в геометрической прогрессии и очень скоро начинают доминировать среди продуктов амплификации.
Амплификатор Stratagene Mx3005Pro оснащён программным обеспечением MxPro™ QPCR, которое работает в операционной системе Microsoft Windows на платформе NT и позволяет:
- осуществлять ввод параметров эксперимента;
- отображать кривые накопления продукта реакции в режиме реального времени;
- устанавливать базовую линию и пороговый цикл автоматически или вручную;
- оценивать абсолютное количество ДНК/РНК при одновременном анализе нескольких калибровочных кривых;
- оптимизировать праймеры и зонды при помощи специального программного модуля;
- осуществлять мониторинг параметров прибора.
Алгоритм проведения ПЦР «в режиме реального времени» можно разделить на стадии:
1) дизайн эксперимента;
2) проведение эксперимента;
3) сбор данных;
4) анализ полученных результатов.
Последовательность выполнения операций:
Последовательность выполнения операций:
– Загрузили операционную систему, включили амплификатор и запустили программное обеспечение с помощью ярлыка «MxPro». После инициализации прибора в появившемся стартовом окне «New Options» выбрали тип анализа исследуемых образцов и нажали кнопку «Ok» (рисунок 2.5). Перед началом работы прогрели галогеновую лампу оптического блока амплификатора, нажав одноименную кнопку на панели инструментов.

Рисунок 2.5 – Стартовое диалоговое окно
– Сформировали схему планшета: расположение образцов в модуле и вид детекции флуоресцентного сигнала с учетом специфики образцов в окне «Plate Setup».

Рисунок 2.6 – Пример заполнения планшета
– Все образцы обозначили значком «Unknown», отметили галочкой имена флуорофоров, которые нужно детектировать, нажали кнопку «Showwellnames» и ввели имя образца в каждую ячейку (рисунок 2.6 и 2.7).

Рисунок 2.7 – Выбор каналов детекции
– Создали программу в модуле «Thermal Profile Setup», с учетом вида анализа и указанием временных температурных режимов, количество циклов. Обязательным шагом является установка этапа считывания флуоресцентного сигнала. Обычно считывание проводят при наименьшем значении температуры в цикле реакции (кроме случаев использования интеркалирующих красителей). Затем сохранили сформированный файл и запустили выполнение программы, нажав кнопку «Run» (рисунок 2.8).

Рисунок 2.8 – Настройка протокола параметров реакции
– Поместили микропробирки или стрипы в реакционный модуль амплификатора и нажали на клавишу «Start». Перед стартом включили (отметили галочкой) опцию выключения лампы после окончания выполнения программы с целью сохранения ресурса лазерной лампы (рисунок 2.9).


Рисунок 2.9 – Окно запуска эксперимента

Рисунок 2.10 – Температурный градиент амплификации
На рисунке 2.10 показан типичный профиль амплификации, выполняемый программным обеспечением амплификатора Stratagene Mx3005Pro.

Рисунок 2.11 – Кривые накопления флуоресцентного сигнала


Рисунок 2.12 – Окно просмотра графиков накопления РНК в одной лунке
При проведении анализа в конце каждого из множества циклов ПЦР регистрируется изменение компонентов реакций, а не после ее завершения. Визуализация регистрации накопления продуктов ПЦР в ходе реакции возможна при нажатии «Show Plots», при этом возможно индивидуально отслеживать интересующую нас ячейку (рисунок 2.12).
На основании считываемых данных управляющая программа строит кривые накопления флуоресцентного сигнала по каждому из заданных для образцов каналов (рисунок 2.13).

Рисунок 2.13 – Результаты амплификации образцов
Результат считается достоверным только в случае прохождения положительных и отрицательных контролей амплификации и отрицательного контроля выделения ДНК.
Рисунок 3.1 – Результат накопления РНК вируса гепатита С в одной лунке
Анализ и учёт результатов всех исследуемых образцов проводили в соответствии следующих правил:
1. Анализируемый образец учитывается как отрицательный (не содержащий РНК ВГС), если для этого образца значение Ct по каналу «ROX» больше 40 или не определяется. Если для такого образца значение Ct ВКО отличается от значения (Ct ВКО)ср более чем на 2, то результат по данному образцу не подлежит анализу и учету как отрицательный. В этом случае необходимо провести повторный анализ данного образца, начиная с этапа выделения (повторный забор крови и повторное проведение анализа).
2. Анализируемый образец считается положительным, т.е. содержащим РНК ВГС, если для этого образца значение Ct по каналу «ROX» меньше или равно 40. Если для такого образца Ct ВКО отличается от значения (Ct ВКО)ср более чем на 2, то такой образец учитывается как положительный без указания концентрации РНК. Для количественного анализа необходимо провести повторный анализ данного образца, начиная с этапа выделения.
3. В случае контаминации все положительные результаты по данной индивидуальной постановке ПЦР считаются недостоверными (требуется ликвидация источника контаминации и повторный анализ всех образцов данной постановки). Образцы, анализ которых дал отрицательный результат, следует учитывать как отрицательные.
4. Если для СПКО значение Ct ВГС больше 40 или не определяется, то все отрицательные результаты считаются недостоверными и требуют перестановки [53].
Для количественного анализа рассчитывали концентрацию РНК ВГС в исследуемых образцах с применением сервисной компьютерной программы «РеалБест Диагностика», которая приводит результаты в виде таблицы с концентрациями РНК ВГС в анализируемых пробах.

Рисунок 3.2 – Программа «РеалБест Диагностика» для количественного учёта результатов концентрации РНК ВГС
В случае отсутствия указанного программного обеспечения или при постановке реакции на приборах количественный анализ специфической вирусной РНК производили по приведённой ниже формуле 3.1:
| СОБРk = CПКО × 2(Ct ПКО – Ct ОБРk) × 2(Ct ВКОk – Ct ОБРПКО),
| (3.1)
|
где k – номер образца;
CПКО – концентрация ПКО, указанная во вкладыше к набору;
Сt ПКО и Ct ВКОПКО – значения Сt данного образца ПКО по каналам ВГВ/ВГС/ВИЧ и FAM, соответственно;
Ct ОБРk и Ct ВКОk – значения Ct данного k-ого образца по каналам «ROX» и «FAM» соответственно.
При условии выделения РНК из образца объёмом 1 мл, полученное значение соответствует концентрации РНК ВГС в МЕ/мл. Если выделение РНК образца производилось из меньшего объёма, полученное значение следует умножить на µ, где µ = 1/объём образца в мл. Таким образом будет получена концентрация РНК ВГС в МЕ/мл. Диапазон определяемых концентраций (область линейности): от 15 МЕ/мл до 108 МЕ/мл РНК ВГС при выделении РНК из 1 мл пробы.
Для определения вирусной нагрузки в копиях РНК ВГС на мл применяли соотношение МЕ = 2,5 копии РНК ВГС (WHO 2nd International Standard for Hepatitis C Assays NIBSC Code: 96/ 798).
Результаты исследований сывороток крови на наличие ВГС, а также качественный и количественный учёт результатов представлены в таблице 1.
Таблица 1. – Результаты исследования сывороток крови на наличие ВГС методом ПЦР «в режиме реального времени»
| № образца п/п
| ПЦР «в режиме реального времени»
|
| Качественное
определение
| Количественное определение
|
| МЕ/мл
| копий/мл
|
| 1.
| +
| 88 200
| 220 500
|
| 2.
| +
| 1 390
| 3 475
|
| 3.
| –
| –
| –
|
| 4.
| –
| –
| –
|
| 5.
| +
|
|
|
| 6.
| –
| –
| –
|
| 7.
| +
| 1120; 970
| 2 425
|
| 8.
| +
| 149 470
| 373 675
|
| 9.
| +
| 31 864
| 79 660
|
| 10.
| –
| –
| –
|
| 11.
| +
| 2 932
| 7 330
|
| 12.
| +
| 10 939
| 27 347
|
| 13.
| +
| 6 699
| 16 747
|
| 14.
| +
| 7 536
| 18 840
|
| 15.
| +
| 2 082 140
| 5 205 350
|
| 16.
| –
| –
| –
|
| 17.
| +
| 2 740
| 6 850
|
| 18.
| +
| 33 217
| 83 043
|
| 19.
| –
| –
| –
|
| 20.
| +
| 23 816
| 59 540
|
| 21.
| +
| 32 943
| 82 357
|
| 22.
| –
| –
| –
|
| 23.
| –
| –
| –
|
|
| –
| –
| –
|
| 25.
| –
| –
| –
|
| 26.
| +
| 16 494
| 41 235
|
| 27.
| –
| –
| –
|
| 28.
| +
|
|
|
| 29.
| –
| –
| –
|
| 30.
| +
| 8 346
| 20 865
|
| 31.
| –
| –
| –
|
| 32.
| –
| –
| –
|
| 33.
| –
| –
| –
|
| 34.
| –
| –
| –
|
| 35.
| –
| –
| –
|
| 36.
| –
| –
| –
|
| 37.
| –
| –
| –
|
ЗАКЛЮЧЕНИЕ
БИБЛИОГРАФИЧЕСКЙ СПИСОК
1. Шляхтенко, Л.И. Пути совершенствования эпидемиологической диагностики вирусных гепатитов В и С / Л.И. Шляхтенко [и др.] // Мир вирусных гепатитов. – 2006. – № 3. – С. 22-25.
2. Ершова, О. В. Современные проявления эпидемического процесса гепатита С, активность естественных путей передачи и совершенствование профилактики этой инфекции: автореф. дисс. докт. мед.наук / О.В. Ершова. – М., 2006. – С 22.
3. Мазепа, В.Н. Оптимизация и комплексное использование полимеразной цепной реакции в диагностике актуальных заболеваний на модели острых кишечных, хеликобакторной, негонококковых, урогенетальных инфекций и вирусных гепатитов: автореф. дисс. док.биол. наук / В.Н. Мазепа. – М. – 2010. – 65 с.
4. Мамедов, М.К. К истории открытия и изучения вируса гепатита С / М.К. Мамедов, М.И. Михайлов // Мир вирусных гепатитов. – 2010. – № 2. – С. 5-8.
5. Мукомолов, Молекулярная эпидемиология вирусных гепатитов / С.Л. Мукомолов, О.В. Калинина // Мир вирусных гепатитов. – 2003. – № 12. – С. 5-13.
6. Шустов, А.В. Генотипическое разнообразие изолятов и молекулярная вариабельность вируса гепатита С у населения Новосибирской области: автореф. дисс. канд. биол. наук / А.В. Шустов. – Кольцово, 2003. – 22 с.
7. Colina, R. Evidence of intratypic recombination in natural populations of hepatitis C virus / R. Colina, D. Casane, S. Vasquez // Gen. Virol. – 2004. – Vol. 85(1). – P. 31-37.
8. Kalinina, O. A natural intergenotypes recombinant of hepatitis C virus identified in St / O. Kalinina, H. Norder, S. Mukomolov // Peterburg.Virol. – 2002. – Vol. 76(8). – P. 4934-4043.
9. Kurbanov, F. Molecular epidemiology and interferon susceptibility of the natural recombinant of hepatitis C virus strain RF1_2k/1b / F. Kurbanov, Y. Tanaka, E. Chub // Infect. Dis. – 2008. – Vol. 198. – P. 1448-1456.
10. Kuiken, C. Los Alamos National Laboratory / C. Kuiken, K. Yusim, L. Boykin, R. Richardson // The Los Alamos HCV Sequence Database. – 2005. – US National Library of Medicine National Institutes of Health.
11. Podzorski, R. Molecular testing in the diagnosis and management of hepatitis C virus infection / R. Podzorski // Arch Pathol Lab Med. – 2002. – Vol. 126(3). – P. 285-290.
12. Suzuki, T. Molecular biology of hepatitis C virus / T. Suzuki, H. Aizaki, K. Murakami // Gastroenterol. – 2007. – Vol. 42. – P. 411-423.
13 Про печенку [Электронный ресурс] / Что такое гепатит С. – Режим доступа: http://propechenku.ru/gepatit/C/chem-opasen.html. – Дата доступа: 11.02.2017
14. Шахгильдян, И.В. Парентеральные вирусные гепатиты (эпидемиология, диагностика, профилактика) / М.И. Михайлов, Г.Г. Онищенко // М.: ГОУ ВУНМЦ МЗ РФ. – 2003. – 384 с.
15. Новикова, Н.А. Вирусология / В.В. Новиков, Н.А. Добротина // Н.Новгород: ННГУ. – 2002. – С. 182-204
16. Быстрова, Т.Н. Парентеральные вирусные гепатиты: этиология, эпидемиология, диагностика, профилактика / Т.Н. Быстрова, Е.И. Ефимов, А.Н. Арзяева, // Учебное пособие для студентов медицинских ВУЗов. – Н.Новгород: НГМА. – 2010. – 180 с.
17. Иванова, Т.Г. Проблемы вирусных гепатитов в современный период / Т.Г. Иванова, Е.М. Яковчук, В.С. Высоцкий // Сборник трудов Всерос. НПК «Современные проблемы эпидемиологии. Перспективные средства и методы лабораторной диагностики и профилактики актуальных инфекций». – СПб. – 2009. – С. 215.
18. Чахарьян, В.В. Особенности эпидемиологии и оценка путей передачи возбудителей вирусных гепатитов В и С в современный период: автореф. дисс. канд. мед.наук / В.В. Чахарьян. – М. – 2009. – 22 с.
19. Ильченко, Л.Ю. Вирус гепатита С и особенности течения ВГС-инфекции у онкогематологических больных / Л.Ю. Ильченко, Р.С. Осканова, Г.И. Сторожаков // Мир вирусных гепатитов. – 2010. – № 2. – С. 9-14.
20. Шерлок, С. Гепатит С (Диагностика, эпидемиология, лечение, профилактика) / С. Шерлок // (Российский консенсус). – Москва. – 26-27 сентября 2000. – № 3(10). – С. 3-92.
21. Thelu, M. Lack of clinical significance of variability in the internal ribosome entry site of hepatitis C virus / M. Thelu, E. Drouet, M. Hilleret // Med. Virol. – 2004. –Vol. 72(3). – P. 396-405.
22. Kunkel, M. Biophysical characterization of hepatitis C virus core protein: implications for interactions within the virus and host / M. Kunkel, SJ. Watowich // FEBS Lett. – 2004. – Vol. 557(1-3). – P. 174-180.
23. Wang, Q. DC-SING: bilding receptors for hepatitis C virus / Z. Feng, Q. Nie // Clin. Med. – 2004. – Vol. 117(9). – P. 1395-1400.
24. Кожанова, Т.В. Лекарственная устойчивость вируса гепатита С (по материалам 45-го Международного конгресса ЕАSL, 14-18 апреля 2010 г.) // Т.В. Кожанова. – Мир вирусных гепатитов. – 2010. – № 2. – С. 32-35.
25. Kamegaya, Y. Hepatitis C virus acts as a tumor accelerator by blocking apoptosis in a mouse model of hepatocarcinogenesis / Y. Kamegaya, Y. Hiasa, L. Zukerberg // Hepatology. – 2005. – Vol. 41(3). – Р. 660-667.
26. Dubuisson, J. Hepaitis C virus proteins / J. Dubuisson // Gastroenterol. – 2007. – Vol. 5. – P. 2406-2415.
27. Moradpour, D. Replication of hepatitis C virus / D. Moradpour, F. Penin, C. Rice // Microbiol. – 2007. – Vol. 5. – P. 453-463.
28. Brun, P. Innate immune system responses differ during recent and chronic hepatitis C virus infection / P. Brun, S. Boninsegna, G. Palu // Hepatology. – 2010. – Vol. 52 (1). – P. 671.
29. ГепПортал [Электронный ресурс] / Вирус гепатита С. – 2010. – Режим доступа: http://www.hv-info.ru/gepatit-s/virus.html. – Дата доступа: 19.02.2017.
30. ГепПортал [Электронный ресурс] / Генотипирование – определение генотипа вируса гепатита С (1, 2, 3, 4). – 2016. – Режим доступа: http://www.hv-info.ru/gepatit-s/analizy/genotipirovanie.html. – Дата доступа: 19.02.2017.
31. Lavillette, D. Characterization of host-range and cell entry properties of the major genotypes and subtypes of hepatitis C virus / D. Lavillette, A. Tarr, C. Voisset // Hepatology. – 2005. – Vol. 41(2). – P. 265-274.
32. Николаева, Л.И. Новые данные о ВГС-инфекции (по материалам 10 Междун. симпозиума по вирусным гепатитам и заболеваниям печени) / Л.И. Николаева // Вектор-Бест. – 2000. – № 3 (17). – C. 3449.
33. Okura, I. Effect of mutation in the hepatitis C virus nonstructural 5B region on HCV replication / I. Okura, N. Horiike, K. Michitaka // Gastroenterol. – 2004. – Vol. 39(5). – P. 449-454.
34. Herrmann, E. Hepatitis C virus kinetics and resistance mechanisms / E. Herrmann, C. Sarrazin // Gastroenterol. – 2004. – Vol. 42 (5). – P. 387-396.
35. Balan, V. Resistance mutation in hepatitis C virus NS5B polymerase gene in patients with chronic hepatitis C virus infection genotype 1b treated with pegylated IFN and ribavirin / V. Balan, A.Pivert, V.Dumbrava // Hepatology. – 2010. – Vol. 52(1). – P. 296.
36. Legrand-Abravanel, F. New natural intergenotypic (2/5) recombinant of hepatitis C virus / F. Legrand-Abravanel, J. Claudinon, F. Nicot // Virol. – 2007. – Vol. 81(8). – P. 4357-4362.
37. Про печенку [Электронный ресурс] / Что такое гепатит С: причины возникновения и виды. – 2015. – Режим доступа: http://propechenku.ru/gepatit/C/chem-opasen.html. – Дата доступа: 24.03.2017.
38. О гепатите.ru [Электронный ресурс] / Особенности устойчивости вируса гепатита С во внешней среде. – 2015. – Режим доступа: http://ogepatite.ru/c/ustojchivost-virusa-gepatita-c-vo-vneshnej-srede.html. – Дата доступа: 27.03.2017.
39. Медицинский портал [Электронный ресурс] / Гепатит С. – Москва. – 2015. – Режим доступа: http://medportal.ru/enc/infection/hepatitis/7/. – Дата доступа: 27.03.2017.
40. Диго, Р.Н. Взаимосвязь серологического профиля при вирусных гепатитах В и С и результатами ПЦР / Р.Н. Диго, А.А. Хорошевская // Сборник трудов НПК с международ. участием «Болезни печени в клинической практике». – Харьков. – 2007. – С. 67-68.
41. Рахманова, А.Г. Вирусные гепатиты (этиопатогенез, эпидемиология, клиника, диагностика и терапия) / А.Г. Рахманова, В.А. Неверов, Г.И. Кирпичникова // Пособие для врачей. – Кольцово. – 2001. – 46 с.
42. Рогозина, Н.В. Клинико-лабораторная характеристика острого вирусного гепатита С у детей / Н.В. Рогозина, А.Л. Мукомолова, Л.Г. Горячева // Педиатрия. – 2004. – № 6. – С. 25-29.
43. Орлов, С.Т. Разработка и апробация тест-системы «АмплиСенс HCV-генотип FRT» на клиническом материале и контрольных панелях QCMD / С.Т. Орлов, А.Д. Неверов, Г.В. Михайловская // Сборник трудов 6-ой всероссийской НПК с международ. участием «Молекулярная диагностика-2007». – М. – 2007. – Т.1. – С. 320-323.
44. Лаптинов, И.А. ПЦР-диагностика без электрофореза. Лабораторная диагностика России / И.А. Лаптинов // Мир медицины: Ежегодный справ. – М. – 2004-2005. – С. 162-163.
45. Чуланов, В.П. Роль молекулярных методов диагностики в оптимизации алгоритмов / В.П. Чуланов, Г.А. Шипулин // Лабораторная медицина. – № 8. – 2006. – С. 67-76.
46. Watkins-Ri






