Сон - физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром. Сон является жизненно необходимым для высших животных и человека. Почему мы останавливаемся подробно именно на этом функциональном состоянии ЦНС? Во-первых, существует много заболеваний, для которых характерны нарушения сна, во-вторых, сон может быть лечебным и профилактическим фактором. Любое функциональное состояние характеризуется по нескольким признакам: изменения на ЭЭГ, мышечный тонус, вегетативные проявления.
Идентификацию сна, его фаз и стадий физиологи проводят на основе общепринятых, так называемых полиграфических, критериев, полиграмм - электроэнцефалограммы (ЭЭГ), электромиограммы (ЭМГ), электроокулограммы (ЭОГ).
Засыпание обычно начинается с появления вместо альфа-ритма, свойственного спокойному бодрствованию (частота 8-12 Гц), тета-волн, (частота 3-7 Гц) это первая стадия сна – дремота.
Вторая стадия характеризуется появлением на ЭЭГ сонных веретен (билатеральные «острые волны») и К-комплексов (частота 12-15 Гц) – эта стадия поверхностного, легкого сна.
В третьей стадии сонные веретена сочетаются с дельта волнами (частота 0.5 – 2 Гц), в третьей стадии дельта волны на ЭЭГ составляют не менее 50%. Это стадия глубокого сна.
Четвертая стадия – на ЭЭГ регистрируются только дельта волны без веретен, для этих волн характерна самая высокая амплитуда и самая низкая частота 0.5 – 2 Гц. В этой стадии наблюдается самый крепкий сон, он так и называется – дельта сон, или медленноволновой - глубокий сон. Как видим, на ЭЭГ фиксируется в это время максимальная амплитуда и минимальная частота, следовательно, максимальная синхронизация активности нейронов.
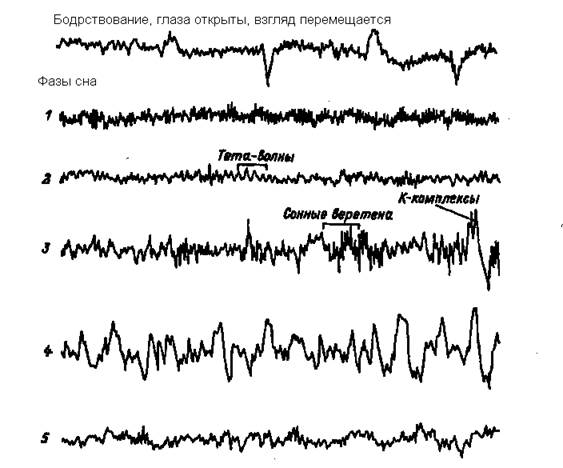
Рисунок 4 ЭЭГ человека в период бодрствования и сна
1 преобладание альфа-ритма, 2 появление тета-волн, 3 появление сонных веретен и К-комплексов, 4 дельта-волны, 5 парадоксальная фаза сна
Именно во время этой фазы сна наблюдаются самые значительные изменения в состоянии организма, характерные для медленноволнового сна: падает тонус скелетных мышц, и человек длительное время может сохранять одно положение тела. Снижается частота сердечных сокращений, артериальное давление, частота дыхания, температура тела, уменьшается секреторная и двигательная активность кишечника и темп метаболизма в целом. При этом ритм дыхания и работы сердца становится более равномерным. Во время этой стадии труднее всего разбудить человека, потому что увеличиваются пороги сенсорной чувствительности. При пробуждении во время самого глубокого сна человек с трудом ориентируется, теряется во времени, долго приходит в себя.
Четвертая стадия сменяется пятой, которая рассматривается отдельно, как совершенно другой вид сна – БДГ сон, потому, что в это время можно наблюдать быстрые движения глаз под закрытыми веками. Эта фаза получила еще и название парадоксального сна. Первый период БДГ, наступающий примерно через девяносто минут после засыпания, - самый короткий, он обычно длится от пяти до десяти минут. По мере продолжения нашего путешествия через ночь длительность каждой последующей фазы БДГ возрастает. Самая длинная из них, которая может занимать более получаса, наступает утром, как раз перед пробуждением. Всего за ночь у здорового взрослого человека при 7-8 часовом сне происходит смена 4 – 6 циклов сна. В первую половину ночи преобладает дельта сон, во вторую происходит удлинение фазы парадоксального сна, длительность дельта сна уменьшается. В норме пробуждение происходит во время фазы парадоксального сна.
Во время парадоксального сна на ЭЭГ фиксируется ритм, характерный для активного бодрствования, вегетативная нервная система, судя по уровню АД, частоте дыхания, уровням некоторых гормонов, переживает настоящую бурю. Повышение активности вегетативной нервной системы выражены настолько, что возможно появление сердечной и дыхательной аритмии. Однако скелетные мышцы практически парализованы, полностью расслаблены, лишь в некоторых мелких мышцах могут наблюдаться подергивания. Именно за это противоречие между картиной активного бодрствования на ЭЭГ и полным параличом скелетной мускулатуры БДГ сон получил название парадоксального. Глазные яблоки совершают быстрые движения под сомкнутыми веками, именно в это время человек видит сны. Если разбудить спящего во время БДГ сна, то в 90% случаев он расскажет о ярком сновидении, причем точность деталей будет существенно выше, чем при пробуждении во время медленного сна. Важной особенностью БДГ сна является еще большее увеличение порогов сенсорной чувствительности. Если во время медленного сна они увеличиваются на 30 – 40%, то во время БДГ сна на 400%, организм практически не воспринимает раздражителей окружающей и внутренней среды.
Парадоксальная стадия сна характерна не только для человека, но и для млекопитающих и птиц. У взрослого человека на БДГ сон приходится 15 – 20% всего сна, у новорожденных до 80%.
На сегодня стало понятно, что необходимым и достаточным признаком сна можно считать ритмичность, т.е. чередование определенных физиологических признаков (полиграфических картин), позволяющих отличить нормальный сон от монотонных "сноподобных состояний". Соответственно и критерием "нормальности" сна служит циклическое чередование стадий 1-2-3-4 медленного сна, которое завершается парадоксальной фазой. На основе такого подхода современное определение сна звучит следующим образом: это "особое генетически детерминированное состояние организма человека (и теплокровных животных, т.е. млекопитающих и птиц), характеризующееся закономерной последовательной сменой определенных полиграфических картин в виде циклов, фаз и стадий".
Что же лежит за этим циклическим чередованием? Каково назначение каждой из двух фаз сна? В физиологии для понимания функций отдельного органа применяют классический метод разрушения: если повредить или удалить данный орган, то, зная последствия и адекватно истолковывая их, можно выяснить его роль. Подобный подход используют и в отношении сна: не давать испытуемому или подопытному животному спать в течение некоторого времени и посмотреть, что при этом меняется в организме и поведении. Впервые такие опыты выполнила более 100 лет назад русская ученая М.М. Манасеина (1843-1903), которая стала основоположником "науки о сне" - сомнологии. В нашем столетии в экспериментах на животных и в наблюдениях за здоровыми людьми неоднократно пытались выяснить, к чему приводит лишение сна (депривация). Однако лишь с использованием электроэнцефалографии такие попытки получили научное обоснование. Исследования последних лет на людях дали до некоторой степени парадоксальные результаты: депривация в течение одних или нескольких суток наиболее мягким, щадящим способом не приводила к серьезным нарушениям в организме и психике субъектов. Наблюдалась лишь повышенная сонливость, утомляемость, раздражительность, рассеянность. Казалось, что главный результат лишения сна - нарастающая потребность в нем! Естественно, подобные работы на людях не могут длиться более 2-3 суток; поэтому последствия длительного лишения сна изучают только в опытах на животных. Так, в 80-е годы группа американских специалистов (А.Речшаффен и сотрудники) получила принципиально важные результаты. Как показали эксперименты, если при первых признаках сна на ЭЭГ (появление сонных веретен и дельта-волн) животных будить, то наступает временное "дробление" сна на очень короткие периоды и его пространственная "локализация", когда сон протекает в отдельных участках мозга. Подобный феномен описан в опытах на обезьянах. Другая группа исследователей наблюдала чередующийся однополушарный медленный сон у дельфинов и ушастых тюленей. Сопоставив эти результаты с некоторыми другими данными по хронической депривации с помощью физических методов, исследователи пришли к неожиданному выводу: полностью исключить медленный сон в принципе невозможно. Как показали эксперименты, через несколько недель от начала хронической депривации у крыс "давление" медленного сна уменьшилось, и если депривация прекращалась, то "отдачи" медленного сна не наблюдалось. Ясно, что вначале это "давление" растет, а затем, по достижении некоторого критического уровня, - спадает на нет в результате постепенной адаптации медленного сна к условиям депривации.
Совершенно противоположные результаты получены в отношении парадоксального сна. Опыты Речшаффена и сотрудников продемонстрировали, что, какой бы вид депривации сна ни проводился (тотальное лишение сна, избирательное лишение медленной или парадоксальной фазы), в результате всегда критично угнетение именно парадоксального сна. Рано или поздно оно приводит к одним и тем же драматическим последствиям (изменению внешнего вида, поведения и внутренних органов), которые через несколько "бессонных" недель завершаются неизбежной гибелью животных. Характерно, однако, что непосредственной причины их гибели обнаружить не удалось. Интересно, что у крыс наблюдалось резкое падение амплитуды ЭЭГ после хронической депривации, которое возникало каждый раз примерно за сутки до гибели животного. Если на этом фоне эксперимент прекращался, то крыса уже не могла заснуть и амплитуда ЭЭГ не восстанавливалась; смерть все равно наступала в течение суток. Следовательно, это падение амплитуды ЭЭГ указывало на какое-то необратимое нарушение работы мозга, вызванное лишением парадоксального сна. Если же опыт прекращался на позднем этапе депривации, но до наступления этого критического момента, то наблюдалась мощная "отдача" только парадоксального сна, независимо от того, какой вид депривации применялся - лишение всего сна, парадоксального или медленного. Таким образом, опыты с длительным лишением сна у лабораторных животных еще раз показывают, что сон включает два принципиально различных состояния организма - медленноволновую и парадоксальную фазы.
Гипотезы происхождения сна.
С физиологическими механизмами сна мы познакомимся, перечислив коротко существующие теории сна. В самом деле, к настоящему времени не существует единой, объясняющей все механизмы, теории сна. Поэтому то, что называется теориями, строго можно назвать лишь гипотезами, однако любая из этих гипотез имеет и экспериментальные, и клинические подтверждения, поэтому каждая из них помогает понять физиологические механизмы сна.
Сон – это одно из функциональных состояний ЦНС. Переход из одного ФС в другое может быть связан с дополнительным сенсорным воздействием, например, возникший звуковой или зрительный сигнал переводит ЦНС из состояния спокойного бодрствования в состояние соответствующее ориентировочному рефлексу. Существовала теория сна, связывающая сон с отсутствием сенсорных раздражителей. Под наблюдением находилась женщина, у которой из всех органов чувств был только один глаз, она засыпала, если этот глаз закрывали. В самом деле, не загруженность аналитической и интегративной деятельностью, монотонная деятельность способствуют развитию сна. Напротив, активная деятельность, шум, яркий свет, боль и другие виды сенсорной афферентации препятствуют развитию сна.
Существует гуморальная теория сна. Эта теория в качестве причины сна рассматривает вещества, появляющиеся в крови при длительном бодрствовании. Гуморальная теория подтверждается экспериментами с перекрестным кровообращением, когда собака, к которой поступает кровь от другой, длительно лишенной сна собаки, засыпает. Эта теория в определенной степени получила подтверждение. В настоящее время выделено уже более десяти пептидов, которые оказывают сомногенный эффект (например пептид дельта сна, СР пептид), структура большинства расшифрована и они даже применяются для регуляции сна. Установлено, что ангиотензин, ТТГ тормозят развитие сна. Вместе с тем, гуморальной теории противоречат наблюдения за сросшимися близнецами, имеющими общее кровообращение: сон одного из близнецов совсем не обязательно приводит к засыпанию другого.
В лаборатории И.П. Павлова было установлено, что при длительной выработке тонкого дифференцировочного торможения животные часто засыпали. Поэтому ученый рассматривал сон как следствие процессов внутреннего торможения, как углубленное, разлитое, распространившееся на оба полушария и ближайшую подкорку торможение (корковая теория сна).
С середины 20 века устойчивые позиции занимают нервные теории сна. Один из главных вопросов, волновавших физиологов еще со времен Павлова, - это существование в мозге "центра сна". Первые указания на существование таких центров связаны с именем Фон Экономо. На рубеже 19 и 20 веков разразилась эпидемия летаргического энцефалита, который протекал либо в виде летаргического сна, либо в виде летаргического бодрствования. Экономо было высказано предположение о существовании двух центров: сна и бодрствования. Поражение одного из них и приводит к наблюдаемому состоянию.
Впервые подтверждено наличие таких центров в экспериментах с электрическим раздражением переднего гипоталамуса (В. Гесс). Такое воздействие приводило к засыпанию.
Расположение центров сна в районе продолговатого и среднего мозга было установлено Ф. Бремером в 1935 году с помощью перерезок мозга на различных уровнях. Было установлено: как только ствол мозга отделяют от больших полушарий, животное (опыты проводились на высших животных) погружается в беспробудный сон. Стало, ясно, что именно тут, в стволе мозга, действует какой-то механизм, организующий ритм сон-бодрствование.
1967 г. Жуве в стволе мозга была открыта новая структура, играющая критическую роль в развитии медленного сна: ядра шва. Разрушение ядер шва устраняло медленный сон. Нейроны ядер шва синтезируют серотонин и направляют его к ретикулярной формации, гипоталамусу, лимбической системе. В латеральной части покрышки ствола мозга обнаружено скопление нейронов, синтезирующих норадреналин (голубое пятно). Стимуляция синего пятна приводит к двигательному возбуждению и изменению ЭЭГ в сторону бодрствования. Между этими двумя системами существуют реципрокные отношения. Серотонин организует медленный сон, НА - бодрствование.
Рядом с синим пятном существует группа гигантских ретикулярных нейронов. Эти нейроны не активны в состояниях бодрствования и медленного сна. Их активность возрастает в период парадоксального сна и подавляется нейронами голубого пятна. Медиатором в этих нейронах является ацетилхолин.
Многие вопросы организации процессов сна получили объяснение с открытием восходящих активирующих влияний ретикулярной формации ствола мозга на кору больших полушарий (Моруцци 1949). Экспериментально было доказано, что сон возникает во всех случаях устранения восходящих активирующих влияний ретикулярной формации на кору мозга. Были установлены нисходящие влияния коры мозга на подкорковые образования. В бодрствующем состоянии при наличии восходящих активирующих влияний ретикулярной формации на кору мозга нейроны лобной коры тормозят активность нейронов центра сна заднего гипоталамуса. В состоянии сна, когда снижаются восходящие активирующие влияния ретикулярной формации на кору мозга, тормозные влияния лобной коры на гипоталамические центры сна снижаются.
Итак, существуют неспецифические активирующие структуры мозга, в частности ретикулярная формация продолговатого и среднего мозга. Частью этой структуры являются ядра шва, голубое пятно и гигантоклеточное ретикулярной ядро, которые и организуют ритм сон-бодрствование с включением парадоксального сна.
Медленный сон и бодрствование можно рассматривать как противоположные состояния. В этом отношении можно согласиться с И.П. Павловым о том, что сон это торможение. К настоящему времени удалось выяснить, что в корковых нейронах действительно наблюдается гиперполяризация, а основном медиатор – ГАМК. Однако и здесь все не так просто. Недавно в опытах на кошках было показано, что по мере развития синхронизации в ЭЭГ первичные нейроны зрительной и слуховой коры перестают реагировать на специфические стимулы и начинают все в большей степени отвечать на импульсацию, приходящую в кору со стороны внутренних органов. Принимая во внимание обнаруженные особые Ca-каналы на мембране многих корковых нейронов, которые открываются при гиперполяризации, можно предположить, что в медленном сне мозг не прекращает перерабатывать информацию, а переходит от обработки внешних сигналов к переработке сигналов от внутренних органов. Таким образом, на пороге XXI в. функция медленного сна, кажется, начинает, наконец, вырисовываться: это восстановление гомеостаза мозговой ткани и оптимизация управления внутренними органами. Для гигиены сна это означает подтверждение старого, как мир, но почему-то забытого правила: без хорошего сна не может быть хорошего бодрствования!
Совершенно по-другому обстоит дело с парадоксальным сном, который, в отличие от медленного сна, имеет ярко выраженную активную природу. На сегодня известно, что парадоксальный сон является обязательной частью сна, центр парадоксального сна расположен в гигантоклеточном ядре ретикулярной формации ствола мозга, этот центр активирует взаимодействие центров сна и бодрствования. Во время парадоксального сна в активирующие системы, использующие в качестве медиаторов ацетилхолин, глутаминовую и аспарагиновую кислоты. Все же остальные активирующие медиаторы (норадреналин, серотонин, дофамин и гистамин) в парадоксальном сне не работают. Это молчание моноаминоэргических нейронов ствола мозга определяет различие между бодрствованием и парадоксальным сном, или на психическом уровне - различие между восприятием внешнего мира и сновидений. Парадоксальный сон жизненно необходим.
Внимание
Рассмотрим теперь физиологические механизмы внимания. Внимание - это сосредоточенность психической деятельности на определенном объекте. С помощью внимания обеспечивается отбор необходимой информации. Как общее состояние сосредоточенности оно связано с повышением возбудимости коры головного мозга в целом или ее отдельных участков. Это в свою очередь соотносится с активностью отдельных частей ретикулярной формации. Те ее отделы, которые своей деятельностью порождают общий эффект возбуждения, входят в структуры, связанные с ориентировочным рефлексом, автоматически возникающим при любых неожиданных и заметных изменениях стимулов, воздействующих на организм. В свою очередь те отделы ретикулярной формации, которые вызывают специфический эффект возбуждения, функционируют, по-видимому, в рамках анатомо-физиологической системы доминанты. С ней же, скорее всего, соотносим в своем действии и избирательный механизм регуляции внимания через актуализацию потребностей, а также механизм волевого управления вниманием через кортикально-подкорковые связи.
Много споров в науке было вокруг вопроса о физиологических основах внимания. Нейрофизиологические механизмы внимания связаны с проявлениями ориентировочного рефлекса, с локальными процессами активации коры головного мозга, а также с эмоциональными состояниями и биологическими потребностями организма. Избирательное внимание возможно лишь при состоянии повышенного, но не чрезмерно высокого бодрствования, которое на ЭЭГ характеризуется высокочастотными десинхронизированными колебаниями низкой амплитуды с редким появлением групп синхронизированных альфа-волн.
Диффузным формам внимания соответствуют состояния расслабленного бодрствования с четко выраженными синхронизированными альфа-ритмами. Рассеянное внимание часто возникает в состоянии сильного возбуждения, при этом на ЭЭГ регистрируется десинхронизированная высокочастотная низкоамплитудная электрическая активность. Анатомическим субстратом регуляции различных форм внимания является ретикулярная формация ствола мозга, диффузная таламическая система, субталамус и гипоталамус. Неспецифическая таламическая система рассматривается как фильтрующий механизм, обладающий способностью переключать внимание с одних раздражителей на другие. Ассоциативные зоны коры являются центральным звеном в системе механизмов, регулирующих отбор информации, избирательные формы восприятия, внимания и сознания.
Непроизвольное внимание заключается в переключении внимания на стимул, который ранее не привлекал внимания. Это переключение происходит автоматически и не требует специальных усилий. Основу непроизвольного внимания, как уже упоминалось, составляет ориентировочный рефлекс. Появление этого рефлекса автоматически включает произвольное внимание и обеспечивает дальнейшую и контролируемую обработку информации.
Ориентировочный рефлекс, или рефлекс «Что такое?» был открыт И.П. Павловым. Каковы же компоненты ориентировочного рефлекса?
1. На уровне коры ориентировочный рефлекс представлен блокадой альфа-ритма и усилением высокочастотных колебаний ЭЭГ.
2. К вегетативным компонентам ориентировочного рефлекса относятся все проявления увеличения симпатической активности: расширение зрачков, изменение ЧСС, периферического сосудистого сопротивления, частоты дыхания, перераспределение кровотока с расширением сосудов головы и сужением периферических сосудов, увеличение кожной проводимости (КГР).
3. На уровне двигательных реакций появление ориентировочного рефлекса выражается в повороте головы и глаз в сторону нового стимула и увеличением мышечного тонуса.
4. Отмечается повышение чувствительности анализаторов.
Отличительной особенностью ориентировочного рефлекса является его угасание, которое развивается при повторении стимула. Произвольное внимание отличается от ориентировочного рефлекса тем, что требует волевых усилий для поддержания уровня активности головного мозга.
Характеристики внимания
Селективность или избирательность внимания характеризуется направленностью на любой аспект стимула – его физическую, лингвистическую характеристику.
Объем внимания определяется количеством одновременно отчетливо осознаваемых объектов. Объем внимания близок объему кратковременной памяти и составляет 7 – 9 стимулов или элементов.
Распределение внимания предполагает распределение его ограниченных ресурсов для одновременного выполнения двух или более заданий.
Устойчивость внимания определяется по длительности выполнения задания, требующего непрерывного внимания.
Под переключением внимания понимают возможность более или менее легкого перехода от одного вида деятельности к другому.
Память






