

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...
Топ:
Проблема типологии научных революций: Глобальные научные революции и типы научной рациональности...
Особенности труда и отдыха в условиях низких температур: К работам при низких температурах на открытом воздухе и в не отапливаемых помещениях допускаются лица не моложе 18 лет, прошедшие...
Интересное:
Принципы управления денежными потоками: одним из методов контроля за состоянием денежной наличности является...
Средства для ингаляционного наркоза: Наркоз наступает в результате вдыхания (ингаляции) средств, которое осуществляют или с помощью маски...
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Учение о нервной системе (неврология)
Общие данные
Одним из основных свойств живого вещества является раздражимость. Каждый живой организм получает раздражения из окружающего его мира и отвечает на них соответствующими реакциями, которые связывают организм с внешней средой. Протекающий в самом организме обмен веществ в свою очередь обусловливает ряд раздражений, на которые организм также реагирует. Связь между участком, на который падает раздражение, и реагирующим органом в высшем многоклеточном организме осуществляется нервной системой.
Проникая своими разветвлениями во все органы и ткани, нервная система связывает все части организма в единое целое, осуществляя его объединение, интеграцию.
Следовательно, нервная система есть "невыразимо сложнейший и тончайший инструмент сношений, связи многочисленных частей организма между собой и организма как сложнейшей системы с бесконечным числом внешних влияний" (И. П. Павлов).
В основе деятельности нервной системы лежит рефлекс (И. М. Сеченов). "Это значит, что в тот или иной рецепторный (воспринимающий. - М. П.) нервный прибор ударяет тот или иной агент внешнего или внутреннего мира организма. Этот удар трансформируется в нервный процесс, в явление нервного возбуждения. Возбуждение по нервным волокнам, как по проводам, бежит в центральную нервную систему и оттуда благодаря установленным связям по другим проводам приносится к рабочему органу, трансформируясь, в свою очередь, в специфический процесс клеток этого органа" (И. П. Павлов).
Основным анатомическим элементом нервной системы является нервная клетка, которая вместе со всеми отходящими от нее отростками носит название нейрона. От тела клетки отходят в одну сторону один длинный (осевоцилиндрический) отросток - аксон или нейрит, в другую сторону - короткие ветвящиеся отростки - дендриты.
|
|
Течение нервного возбуждения внутри нейрона идет в направлении от дендритов к телу клетки и от нее к аксону; аксоны проводят возбуждение в направлении от тела клетки. Передача нервного импульса с одного нейрона на другой осуществляется посредством особым образом построенных концевых аппаратов или синапсов (synapsis греч. - соединение). Различают аксосоматические связи нейронов, при которых разветвления одного нейрона подходят к телу клетки другого нейрона, и филогенетически более новые аксо-дендритические связи, когда контакт осуществляется дендритами нервных клеток (С. А. Саркисов).
Аксо-дендритические связи сильно развиты в филогенетически новых и высших в функциональном отношении верхних слоях коры. Они играют роль в механизме перераспределения нервных импульсов в коре и представляют, по-видимому, морфологическую основу временных связей при условно-рефлекторной деятельности. В спинном мозгу и подкорковых образованиях превалируют аксосоматические связи.
Прерывистость пути нервного проведения выражена повсюду, создавая возможность самых разнообразных связей (Ф. А. Поемный и Е. П. Семенова).
Таким образом, вся нервная система представляет собой комплекс нейронов, которые, вступая в соединение друг с другом, нигде не срастаются непосредственно между собой.
Следовательно, нервное возбуждение, возникнув в каком-либо месте, передается по отросткам нервных клеток от одного нейрона к другому, от другого к третьему и т. д. Наглядным примером связи между органами, устанавливаемой при посредстве нейронов, может служить так называемая рефлекторная дуга, лежащая в основе рефлекса, наиболее простой и вместе с тем самой основной реакции нервной системы.
Простая рефлекторная дуга состоит по крайней мере из двух нейронов, из которых один связан с какой-нибудь чувствительной поверхностью (например, кожей), а другой с помощью своего нейрита оканчивается в мышце (или железе). При раздражении чувствительной поверхности возбуждение идет по связанному с ней нейрону в центростремительном направлении (центрипетально) к рефлекторному центру, где находится соединение (синапс) обоих нейронов. Здесь возбуждение переходит на другой нейрон и идет уже центробежно (центрифугально) к мышце или железе. В результате происходит сокращение мышцы или изменение секреции железы. Часто в состав простой рефлекторной дуги входит третий вставочный нейрон, который служит передаточной станцией с чувствительного пути на двигательный. Кроме простой (трехчленной) рефлекторной дуги, имеются сложноустроенные многонейронные рефлекторные дуги, проходящие через разные уровни головного мозга, включая его кору. У высших животных и человека на фоне простых и сложных рефлексов также при посредстве нейронов образуются временные рефлекторные связи высшего порядка, известные в настоящее время под названием условных рефлексов (И. П. Павлов).
|
|
Таким образом, всю нервную систему можно себе представить состоящей в функциональном отношении из трех родов элементов (рис. 264).

Рис. 264. Схема рефлекторной дуги. 1 - нервное окончание чувствительного нейрона в коже; 2 - периферический отросток чувствительного нейрона; 3 - спинномозговой узел; 4 - центральный отросток чувствительного нейрона; 5 - вставочный нейрон; 6 - двигательная клетка переднего рога; 7 - нейрит двигательной клетки; 8 - нервное окончание в мышце
1. Рецептор (восприниматель), трансформирующий энергию внешнего раздражения в нервный процесс; он связан с афферентным (центростремительным, или рецепторным) нейроном, распространяющим начавшееся возбуждение (нервный импульс) к центру; с этого явления начинается анализ (И. П. Павлов).
2. Кондуктор (проводник), вставочный, или ассоциативный, нейрон, осуществляющий замыкание, т. е. переключение возбуждения с центростремительного нейрона на центробежный и превращение полученного центром импульса во внешнюю реакцию. Это явление есть синтез, который представляет, "очевидно, явление нервного замыкания" (И. П. Павлов). Поэтому И. П. Павлов называет этот нейрон контактором, замыкателем.
|
|
3. Эфферентный (центробежный) нейрон, осуществляющий ответную реакцию (двигательную или секреторную) благодаря проведению нервного возбуждения от центра к периферии, к эффектору - производителю эффекта, действия, т. е. к рабочему органу (мышца, железа). Поэтому этот нейрон называют также эффекторным. Рецепторы возбуждаются со стороны трех чувствительных поверхностей, или рецепторных полей, организма: 1) с наружной кожной поверхности тела (экстероцептивного поля) при посредстве связанных с ней генетически органов чувств, получающих раздражение из внешней среды: 2) с внутренней поверхности тела (интероцептивного поля), принимающей раздражения главным образом со стороны химических веществ, поступающих в полости внутренностей, и 3) из толщи стенок собственно тела (проприоцептивного поля), в которых заложены кости, мышцы и другие органы, производящие раздражения, воспринимаемые специальными рецепторами. Рецепторы от названных полей связаны с афферентными нейронами, которые достигают центра и там переключаются при посредстве подчас весьма сложной системы кондукторов на различные эфферентные проводники; последние, соединяясь с рабочими органами, дают тот или иной эффект.
Общая характеристика нервной системы с точки зрения кибернетики заключается в следующем.
Как уже говорилось, кибернетика учит, что живой организм - это уникальная кибернетическая машина, способная к самоуправлению. Эту функцию выполняет нервная система. Для самоуправления требуются 3 звена: I звено - поступление информации, которое происходит по определенному вводному каналу информации и совершается следующим образом:
А. Возникающее из источника информации сообщение поступает на приемный конец канала информации - рецептор. Рецептор - это кодирующее устройство, которое воспринимает сообщение и перерабатывает его в сигнал - афферентный сигнал в результате чего внешнее раздражение превращается в нервный импульс.
Б. Афферентный сигнал передается далее по каналу информации, каковым является афферентный нерв.
Имеются 3 вида каналов информации, 3 входа в них: внешние входы - через органы чувств (экстероцепторы); внутренние входы - а) через органы растительной жизни (внутренности) - интероцепторы; б) через органы животной жизни (сома, собственно тело) - проприоцепторы. II звено - переработка информации. Она совершается декодирующим устройством, которое составляют клеточные тела афферентных нейронов нервных узлов и нервные клетки серого вещества спинного мозга, коры и подкорки головного мозга, образующие нервную сеть серого вещества центральной нервной системы. III звено - управление. Оно достигается передачей эфферентных сигналов из серого вещества спинного и головного мозга на исполнительный орган и осуществляется по эфферентным каналам, т. е. по эфферентным нервам с эффектором на конце.
|
|
Имеются 2 рода исполнительных органов:
1. Исполнительные органы животной жизни - поперечнополосатые мышцы, преимущественно скелетные.
2. Исполнительные органы растительной жизни - гладкие мышцы и железы.
Кроме этой кибернетической схемы, современная кибернетика установила общность принципа обратной связи для управления и координации процессов, совершающихся как в современных автоматах, так и в живых организмах; с этой точки зрения в нервной системе можно различать обратную связь рабочего органа с нервными центрами, так называемую "обратную афферентацию" (П. К. Анохин). Под этим названием подразумевается передача сигналов с рабочего органа в центральную нервную систему о результатах его работы в каждый данный момент. Когда центры нервной системы посылают эфферентные импульсы в исполнительный орган, то в последнем возникает определенный рабочий эффект (движение, секреция). Этот эффект побуждает в исполнительном органе нервные (чувствительные) импульсы, которые по афферентным путям поступают обратно в спинной и головной мозг и сигнализируют о выполнении рабочим органом определенного действия в данный момент. Это и составляет сущность "обратной афферентации", которая, образно говоря, есть доклад центру о выполнении приказа на периферии. Так, при взятии рукой предмета глаза непрерывно измеряют расстояние между рукой и целью и свою информацию посылают в виде афферентных сигналов в мозг. В мозгу происходит замыкание на эфферентные нейроны, которые передают двигательные импульсы в мышцы руки, производящие необходимые для взятия ею предмета действия. Мышцы одновременно воздействуют на находящиеся в них рецепторы, беспрерывно посылающие мозгу чувствительные сигналы, информирующие о положении руки в каждый данный момент. Такая двусторонняя сигнализация по цепям рефлексов продолжается до тех пор, пока расстояние между кистью руки и предметом не будет равно нулю, т. е. пока рука не возьмет предмет.
|
|
Следовательно, все время совершается самопроверка работы органа, возможная благодаря механизму "обратной афферентации", который имеет характер замкнутого круга в последовательности: центр (прибор, задающий программу действия) - эффектор (мотор) - объект (рабочий орган) - рецептор (восприемник) - центр.
Существование такой замкнутой кольцевой, или круговой, цепи рефлексов центральной нервной системы и обеспечивает все сложнейшие коррекции протекающих в организме процессов при любых изменениях внутренних и внешних условий (В. Д. Моисеев, 1960). Без механизмов обратной связи живые организмы не смогли бы разумно приспособиться к окружающей среде.
Следовательно, вместо прежнего представления о том, что в основе строения и функции нервной системы лежит разомкнутая рефлекторная дуга, теория информации и обратной связи ("обратной афферентации") дает новое представление о замкнутой кольцевой цепи рефлексов, о круговой системе эфферентно-афферентной сигнализации. Не разомкнутая дуга, а сомкнутый круг - таково новейшее представление о строении и функции нервной системы.
Таким образом, в свете данных кибернетики нервная система характеризуется как система информации и управления.
Единая нервная система человека условно делится на 2 части соответственно двум основным частям организма - растительной и животной: 1) часть нервной системы, иннервирующая все внутренности, а также эндокринную систему и гладкие мышцы кожи, сердце и сосуды, т. е. органы растительной жизни, создающие внутреннюю среду организма, называется растительной нервной системой, вегетативной;* 2) другая часть нервной системы, управляющая поперечнополосатой мускулатурой скелета и некоторых внутренностей (язык, гортань, глотка) и иннервирующая главным образом органы животной жизни, называется животной нервной системой, анимальной. Ее также не совсем удачно называют соматической, имея в виду сому, т. е. собственно тело. Она заведует по преимуществу функциями связи организма с внешней средой, обусловливая чувствительность организма (при посредстве органов чувств) и движения мускулатуры скелета. Условность и ограниченность приведенной выше классификации явствует из того, что вегетативная нервная система имеет отношение к иннервации всех органов, в том числе и соматических, так как она участвует в их питании (трофике), а также определяет тонус скелетной мускулатуры.
* (Вегетативную нервную систему иначе называют автономной, хотя известную автономию имеет и спинной мозг, а также висцеральной, но вегетативная иннервация не ограничивается только внутренностями. Также нелогичны такие имеющиеся в литературе обозначения анимальной нервной системы, как цереброспинальная (ибо этот термин относится ко всей нервной системе) и соматическая (ибо сома означает тело вообще, в целом). Все эти неточные термины постепенно выводятся из употребления.)
И. П. Павлов и особенно К. М. Быков со своими учениками (В. Н. Черниговский и др.) показали зависимость деятельности всех внутренностей и сосудов от коры головного мозга,
Вегетативная часть нервной системы в свою очередь делится на два отдела: симпатический и парасимпатический, которые для краткости также называются системами. Симпатическая система иннервирует все тело, а парасимпатическая - лишь определенные области его.
Кроме такой классификации, соответствующей строению организма, нервную систему делят по топографическому принципу на центральный и периферический отделы, или системы. Под центральной нервной системой разумеется спинной и головной мозг, которые состоят из серого и белого вещества, под периферической - все остальное, т. е. нервные корешки, узлы, сплетения, нервы и периферические нервные окончания. Серое вещество спинного и головного мозга - это скопления нервных клеток вместе с ближайшими разветвлениями их отростков, называемые нервными центрами. Нервный центр - это "скопление и сцепление нервных клеток" (И. П. Павлов).
Белое вещество - это нервные волокна (отростки нервных клеток, нейриты), покрытые миелиновой оболочкой (откуда и происходит белый цвет) и связывающие отдельные центры между собой, т. е. проводящие пути. Как в центральном, так и в периферическом отделах нервной системы содержатся элементы анимальной и вегетативной частей ее, чем достигается единство всей нервной системы.
Высшим отделом ее, который ведает всеми процессами организма, как животными, так и растительными, является кора большого мозга.
Развитие спинного мозга. Как уже говорилось выше, филогенетически спинной мозг (туловищный мозг ланцетника) появляется на III этапе развития нервной системы (трубчатой нервной системы). В это время головного мозга еще нет, поэтому туловищный мозг имеет центры для управления всеми процессами организма, как вегетативными, так и анимальными (висцеральные и соматические центры). Соответственно сегментарному строению тела туловищный мозг имеет сегментарное строение, он состоит из связанных между собой невромеров, в пределах которых замыкается простейшая рефлекторная дуга. Метамерное строение спинного мозга сохраняется и у человека, чем и обусловливается наличие у него коротких рефлекторных дуг.
С появлением головного мозга (этап цефализации) в нем возникают высшие центры управления всем организмом, а спинной мозг попадает в подчиненное положение. Спинной мозг не остается только сегментарным аппаратом, а становится и проводником импульсов от периферии к головному мозгу и обратно, и в нем развиваются двусторонние связи с головным мозгом. Таким образом, в процессе эволюции спинного мозга образуются два аппарата: один, более старый, сегментарный аппарат собственных связей спинного мозга и второй, более новый, надсегментарный аппарат двусторонних проводящих путей к головному мозгу. Такой принцип строения наблюдается и у человека.
Решающим фактором образования туловищного мозга является приспособление к окружающей среде при помощи движения. Поэтому строение спинного мозга отражает способ передвижения животного. Так, например, у пресмыкающихся, не имеющих конечностей и передвигающихся с помощью туловища (например, у змеи), спинной мозг развит равномерно на всем протяжении и не имеет утолщений. У животных, пользующихся конечностями, возникает два утолщения; при этом, если более развиты передние конечности (например, крылья птиц), то преобладает переднее (шейное) утолщение спинного мозга; если более развиты задние конечности (например, ноги страуса), то увеличено заднее (поясничное) утолщение; если в ходьбе одинаково участвуют и передние, и задние конечности (четвероногие млекопитающие), то одинаково развиты оба утолщения. У человека в связи с более сложной деятельностью руки как органа труда шейное утолщение спинного мозга дифференцировалось сильнее, нежели поясничное.
Отмеченные факторы филогенеза играют роль в развитии спинного мозга и в онтогенезе. Спинной мозг развивается из мозговой трубки, из ее заднего отрезка (из переднего возникает головной мозг.). Из вентрального отдела мозговой трубки образуются передние столбы серого вещества спинного мозга (клеточные тела двигательных нейронов), прилегающие к ним пучки продольных нервных волокон и отростки названных нейронов (двигательные корешки). Из дорсального отдела возникают задние столбы серого вещества (клеточные тела чувствительных нейронов), задние канатики (отростки чувствительных нейронов).
Таким образом, вентральная часть мозговой трубки является первично двигательной, а дорсальная - первично чувствительной. Деление на моторную (двигательную) и сенсорную (чувствительную) области простирается на всю мозговую трубку и сохраняется в стволе головного мозга.
Из-за редукции каудальной части спинного мозга получается тонкий тяж из нервной ткани, будущая Шит terminate. Первоначально, на 3-м месяце утробной жизни, спинной мозг занимает весь позвоночный канал, затем позвоночник начинает расти скорее, чем мозг, вследствие чего конец последнего постепенно перемещается кверху (краниально). При рождении конец спинного мозга уже находится на уровне III поясничного позвонка, а у взрослого достигает высоты I-II поясничного позвонка. Благодаря такому "восхождению" спинного мозга отходящие от него нервные корешки принимают косое направление (рис. 267).

Рис. 267. Спинной мозг. а - вид спереди (вентральная его поверхность); б - вид сзади (дорсальная его поверхность). Твердая и паутинная оболочки разрезаны. Сосудистая оболочка снята. Римскими цифрами обозначен порядок расположения шейных (С), грудных (Th), поясничных (L) и крестцовых (S) спинномозговых нервов. 1 - intumescentia cervicalis; 2 - ganglion spinale; 3 - dura mater medullae; spinalis; 4 - intumescentia lumbalis; 5 - conns medullaris 6 - cauda equina
Строение спинного мозга
Спинной мозг, medulla spinalis (греч. myelos) лежит в позвоночном канале и у взрослых представляет собой длинный (45 см у мужчин и 41-42 см у женщин), несколько сплюснутый спереди назад цилиндрический тяж, который вверху (краниально) непосредственно переходит в продолговатый мозг, а внизу (каудально) оканчивается коническим заострением, conus medullaris, на уровне II поясничного позвонка (см. рис. 267). Знание этого факта имеет практическое значение (чтобы не повредить спинной мозг при поясничном проколе с целью взятия спинномозговой жидкости или с целью спинномозговой анестезии, надо вводить иглу шприца между остистыми отростками III и IV поясничных позвонков).
От conus medullaris отходит книзу так называемая концевая нить, filum terminate, представляющая атрофированную нижнюю часть спинного мозга, которая внизу состоит из продолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку.
Спинной мозг на своем протяжении имеет два утолщения, соответствующих корешкам нервов верхней и нижней конечностей: верхнее из них называется шейным утолщением, intumescentia cervicalis, а нижнее - поясничным, intumescentia lumbalis. Из этих утолщений более обширно поясничное, но более дифференцировано шейное, что связано с более сложной иннервацией руки как органа труда. Образовавшимися вследствие утолщения боковых стенок спинномозговой трубки и проходящими по средней линии передней и задней продольными бороздками, глубокой - flssura medidna anterior, и поверхностной - sulcus medidnus posterior, спинной мозг делится на две симметричные половины - правую и левую; каждая из них в свою очередь имеет слабо выраженную продольную бороздку, идущую по линии входа задних корешков. Она называется sulcus lateralis posterior.
Названная бороздка и место выхода передних корешков из мозга подразделяют каждую половину спинного мозга на три продольных канатика: передний - funiculus anterior, боковой - funiculus lateralis и задний - funiculus posterior. Задний канатик в шейном и верхнегрудном отделах делится еще промежуточной бороздкой - sulcus intermedius posterior, на два пучка: fasciculus gracilis (пучок Голля) и fasciculus cuneatus (пучок Бурдаха). Оба эти пучка под теми же названиями переходят вверху на заднюю сторону продолговатого мозга.
На той и другой стороне из спинного мозга выходят двумя продольными рядами корешки спинномозговых нервов. Передний корешок, radix ventralis s. anterior, состоит из нейритов двигательных (центробежных или эфферентных) нейронов, клеточные тела которых лежат в спинном мозгу, тогда как задний корешок, radix dorsalis s. posterior, входящий в sulcus lateralis posterior, содержит отростки чувствительных (центростремительных или афферентных) нейронов, тела которых лежат в спинномозговых (межпозвонковых) узлах*.
* (По последним данным (Гассер, 1955; Гесс, 1956; Митчелл, 1957, и др.), в задних корешках проходят безмякотные эфферентные волокна, иннервирующие гладкие мышцы внутренностей и сосудов.)
На некотором расстоянии от спинного мозга двигательный корешок прилегает к чувствительному (рис. 268), и они вместе образуют ствол спинномозгового нерва, который невропатологи выделяют под именем канатика, funiculus. При воспалении канатика (фуникулите) возникают сегментарные расстройства одновременно двигательной и чувствительной сферы; при заболевании корешка (радикулите) наблюдаются сегментарные нарушения одной сферы - или чувствительной, или двигательной, а при воспалении ветвей нерва (неврите) расстройства соответствуют зоне распространения данного нерва. Канатик обычно очень короткий, так как он быстро выходит из межпозвонкового отверстия, после чего спинномозговой нерв распадается на свои основные ветви.

Рис. 268. Элементы периферической нервной системы (схема). 1 - radix dorsalis; 2 - radix ventralis; 3 - ganglion spinale; 4 - funiculus; 5 - plexus; 6 - ветви сплетения; 7 - задний рог; 8 - передний рог
В межпозвонковых отверстиях вблизи места соединения обоих корешков задний корешок имеет утолщение - спинномозговой или межпозвонковый узел, ganglion spinale s. intervertebral, содержащий ложноуниполярные нервные клетки (афферентные нейроны) с одним отростком, который делится затем на две ветви: одна из них, центральная, идет в составе заднего корешка в спинной мозг, другая, периферическая, продолжается в спинномозговой нерв. Таким образом, в спинномозговых узлах отсутствуют синапсы, так как здесь лежат клеточные тела только афферентных нейронов. Этим названные узлы отличаются от вегетативных узлов периферической нервной системы, так как в последних вступают в контакты вставочные и эфферентные нейроны*. Спинномозговые узлы крестцовых корешков лежат внутри крестцового канала, а узел копчикового корешка - внутри мешка твердой мозговой оболочки.
* (В составе спинальных ганглиев могут встретиться мультиполярные элементы вегетативной нервной системы.)
Вследствие того, что спинной мозг короче позвоночного канала, место выхода нервных корешков не соответствует уровню межпозвонковых отверстий. Чтобы попасть в последние, корешки направляются не только в стороны от мозга, но еще и вниз и при этом тем отвеснее, чем ниже они отходят от спинного мозга. В поясничной части последнего нервные корешки спускаются к соответствующим межпозвонковым отверстиям параллельно filum terminate, облекая ее conus medullaris густым пучком, который носит название конского хвоста, cauda equina (см. рис. 267).
Внутреннее строение спинного мозга. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого вещества, слагающегося из миелиновых нервных волокон.
А. Серое вещество, substantia grisea, развившееся из мозговой трубки, из среднего слоя эпителиальных клеток ее, заложено внутри спинного мозга и окружено со всех сторон белым веществом. Серое вещество образует две вертикальные колонны, помещенные в правой и левой половине спинного мозга. В середине его заложен узкий центральный канал, canalis centralis, спинного мозга, проходящий во всю длину последнего и содержащий спинномозговую жидкость. Центральный канал является остатком полости первичной нервной трубки. Поэтому вверху он сообщается с IV желудочком головного мозга, а внизу, в области conus medullaris, заканчивается небольшим расширением - концевым желудочком, ventriculus terminalis. С возрастом центральный канал суживается и местами совсем зарастает, так что после 40 лет (в 93%) он перестает быть сплошным каналом (В. П. Курковский).
Серое вещество, окружающее центральный канал, носит название промежуточного вещества, substantia intermedia centralis. В каждой колонне серого вещества различают два столба: передний, columna grisea anterior, и задний, columna grisea posterior.
На поперечных разрезах спинного мозга эти столбы имеют вид рогов: переднего, расширенного, cornu anterius, и заднего, заостренного, cornu posterius. Благодаря этому общий вид серого вещества, выделяющегося на фоне белого, напоминает букву "Н".
Серое вещество состоит из нервных клеток, группирующихся в ядра, расположение которых в основном соответствует сегментарному строению спинного мозга и его первичной трехчленной рефлекторной дуге. Первый, чувствительный нейрон этой дуги лежит в спинномозговых узлах, периферический отросток его идет в составе нервов к органам и тканям и вступает там в связь с рецепторами, а центральный в составе задних чувствительных корешков проникает через sulcus lateralis posterior в спинной мозг, где вступает в связь с клетками задних рогов. Благодаря этому вокруг верхушки заднего рога образуется пограничная зона белого вещества, представляющая собою совокупность центральных отростков клеток спинальных ганглиев, заканчивающихся в спинном мозгу. Клетки задних рогов образуют отдельные группы или ядра, воспринимающие из "сомы" различные виды чувствительности (кожной и органов движения), - соматически-чувствительные ядра.
Среди них выделяются: ядро основания заднего рога, nucleus thoracicus (столб Кларка - Штиллинга), наиболее выраженное в грудных сегментах мозга, находящееся на верхушке рога студенистое вещество, substantia gelatinosa, а также так называемые собственные ядра - nuclei proprii.
Заложенные в заднем роге клетки образуют вторые, вставочные, нейроны; они дают начало нейритам, идущим в головной мозг, а клетки студенистого вещества и диффузно разбросанные в сером веществе рассеянные клетки, так называемые пучковые клетки, служат для связи с третьими нейронами, заложенными в передних рогах того же сегмента. Отростки этих клеток, идущие от задних рогов к передним, естественно, располагаются вблизи серого вещества, по его периферии, образуя узкую кайму белого вещества, непосредственно окружающего серое со всех сторон. Это собственные, или основные, пучки спинного мозга, fasciculi proprii. Аксоны других пучковых клеток делятся на восходящую и нисходящую ветви, которые оканчиваются на клетках передних рогов нескольких выше и нижележащих сегментов. Вследствие этого раздражение, идущее из определенной области тела, может передаваться не только на соответствующий ей сегмент спинного мозга, но захватывать и другие. В результате простой рефлекс может вовлекать в ответную реакцию целую группу мышц, обеспечивая сложное координированное движение, остающееся, однако, безусловно-рефлекторным.
Передние рога содержат третьи, двигательные, нейроны, аксоны которых, выходя из спинного мозга, составляют передние, двигательные корешки. Эти клетки образуют ядра эфферентных соматических нервов, иннервирующие скелетную мускулатуру - соматически-двигательные ядра*. Последние имеют вид коротких колонок и лежат в виде двух групп - медиальной и латеральной. Медиальные иннервируют мышцы, развившиеся из дорсальной части миотомов (аутохтонная мускулатура спины), а латеральные - мышцы, происходящие из вентральной части миотомов (вентролатеральные мышцы туловища и мышцы конечностей). При этом чем дистальнее находятся иннервируемые мышцы, тем латеральнее лежат иннервирующие их клетки.
* (Клетки передних рогов являются также и трофическими центрами мускулатуры; выключение двигательных нейронов влечет за собой не только паралич, но и атрофию мышц.)
Наибольшее число ядер содержится в передних рогах шейного утолщения спинного мозга, откуда иннервируются верхние конечности, что определяется участием последних в трудовой деятельности человека. У последнего в связи с усложнением движений руки как органа труда этих ядер значительно больше, чем у животных, включая антропоидов. Таким образом, задние и передние рога серого вещества имеют отношение к иннервации органов животной жизни, особенно аппарата движения, в связи с усовершенствованием которого в процессе эволюции и развивался спинной мозг.
Передний и задний рога в каждой половине спинного мозга связаны между собой промежуточной зоной серого вещества, которая в грудном и поясничном отделах спинного мозга, на протяжении от I грудного до II-III поясничных сегментов, особенно выражена и выступает в виде бокового рога, cornu laterale. Вследствие этого в названных отделах серое вещество на поперечном разрезе приобретает вид бабочки. В боковых рогах заложены клетки, иннервирующие вегетативные органы и группирующиеся в ядро, которое носит название nucleus intermediolateralis (впервые описано И. М. Якубовичем). Нейриты клеток этого ядра выходят из спинного мозга в составе передних корешков.
Б. Белое вещество, substantia alba, спинного мозга состоит из нервных отростков, которые составляют три системы нервных волокон:
1. Короткие пучки ассоциационных волокон, соединяющих участки спинного мозга на различных уровнях (афферентные и вставочные нейроны).
2. Длинные центростремительные (чувствительные, афферентные) волокна.
3. Длинные центробежные (двигательные, эфферентные) волокна.
Первая система (коротких волокон) относится к собственному аппарату спинного мозга, а остальные две системы (длинных волокон) составляют проводниковый аппарат двусторонних связей с головным мозгом.
Собственный аппарат включает серое вещество спинного мозга с задними и передними корешками и собственными основными пучками белого вещества (fasciculi proprii), окаймляющими серое в виде узкой полосы. По развитию собственный аппарат является образованием филогенетически более старым и потому сохраняет примитивные черты строения - сегментарность, отчего его называют также сегментарным аппаратом спинного мозга в отличие от остального несегментированного аппарата двусторонних связей с головным мозгом.
Таким образом, нервный сегмент - это поперечный отрезок спинного мозга и связанных с ним правого и левого спинномозговых нервов, развившихся из одного невротома (невромера). Он состоит из горизонтального слоя белого и серого вещества (задние, передние и боковые рога), содержащего нейроны, отростки которых проходят в одном парном (правом и левом) спинномозговом нерве и его корешках (см. рис. 268). В спинном мозге различают 31 сегмент, которые топографически делятся на 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый. В пределах нервного сегмента замыкается короткая рефлекторная дуга (см. рис. 264).
Так как собственный сегментарный аппарат спинного мозга возник тогда, когда еще не было головного, то функция его - это осуществление тех реакций в ответ на внешнее и внутреннее раздражения, которые в процессе эволюции возникли раньше, т. е. врожденных реакций, или безусловных рефлексов.
Аппарат двусторонних связей с головным мозгом является филогенетически более молодым, так как он возник лишь тогда, когда появился головной мозг.
По мере развития последнего разрастались кнаружи и проводящие пути, связывающие спинной мозг с головным (рис. 269). Этим объясняется тот факт, что белое вещество спинного мозга как бы окружило со всех сторон серое вещество. Благодаря проводниковому аппарату собственный аппарат спинного мозга связан с аппаратом головного мозга, который объединяет работу всей нервной системы. Нервные волокна группируются в пучки, различимые на препарате лишь с помощью особых методов (см. курс микроскопической анатомии), а из пучков составляются видимые невооруженным глазом канатики: задний, боковой и передний. В заднем канатике (рис. 270), прилежащем к заднему (чувствительному) рогу, лежат пучки восходящих нервных волокон; в переднем канатике, прилежащем к переднему (двигательному) рогу, лежат пучки нисходящих нервных волокон; наконец, в боковом канатике находятся и те и другие. Кроме канатиков, белое вещество находится в белой спайке, comissura alba, образующейся вследствие перекреста волокон спереди от substantia intermedia centralis; сзади белая спайка отсутствует.
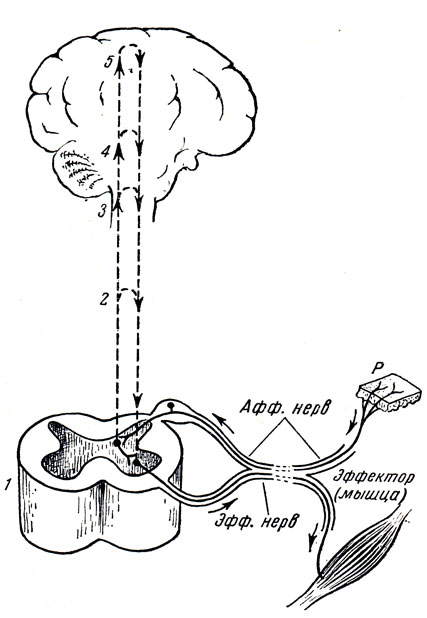
Рис. 269. Элементарная схема безусловного рефлекса. Нервные импульсы, возникающие при раздражении рецептора (Р), по афферентным волокнам (Афф. нерв, показано лишь одно такое волокно) идут к спинному мозгу (1), где через вставочный нейрон передаются на эфферентные волокна (Эфф. нерв), по которым доходят до эффектора. Пунктирные линии - распространение возбуждения от низших отделов центральной нервной системы на ее вышерасположенные отделы (2, 3, 4), до коры мозга (5) включительно. Наступающее вследствие этого изменение состояния высших отделов мозга в свою очередь воздействует (см. стрелки) на эфферентный нейрон, влияя на конечный результат рефлекторного ответа

Рис. 270. а - схема проводящих путей спинного мозга на поперечном разрезе. Слева изображено местоположение восходящих систем волокон, справа - нисходящих систем волокон; 1 - fasciculus gracilis (Goll); 2 - fasciculus cuneatus (Burdagh); 3 - radix posterior; 4 - tractus corticospinalis {lateralis; 5 - tractus rubrospinalis; 6 - tractus tectospinalis; 7 - tractus spinothalamicus lateralis; 8 - tractus spinotectalis; 9 - tractus vestibulospinalis; 10 - tractus olivospinalis; 11 - tractus reticulospinalis; 12 - tractus corticospinalis anterior; 13 - tractus spinocerebellars anterior (Gowers); 14 - tractus spinocerebellars posterior (Flechsig); 15 - fasciculi proprii; 16 - tractus spinothalamicus anterior; 17 - tractus thalamospinalis; б - поперечное сечение спинного мозга; ядра серого вещества (в грудном отделе); 1 - substantia gelatinosa (Rolandi); 2 - nucleus proprius cornu post.; 3 - nucleus thoracicus (столб Кларка - Штиллинга); 4 - nucleus intermediomedialls; 5 - nucleus intermediolateralis; 6, 7, 8, 9, 10 - пять двигательных ядер переднего рога; I, II, III - передний, боковой и задний канатики белого вещества
Задние канатики содержат волокна задних спинномозговых кор
|
|
|

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...

Своеобразие русской архитектуры: Основной материал – дерево – быстрота постройки, но недолговечность и необходимость деления...

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!