

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...

Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...
Топ:
Эволюция кровеносной системы позвоночных животных: Биологическая эволюция – необратимый процесс исторического развития живой природы...
Характеристика АТП и сварочно-жестяницкого участка: Транспорт в настоящее время является одной из важнейших отраслей народного хозяйства...
Техника безопасности при работе на пароконвектомате: К обслуживанию пароконвектомата допускаются лица, прошедшие технический минимум по эксплуатации оборудования...
Интересное:
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Средства для ингаляционного наркоза: Наркоз наступает в результате вдыхания (ингаляции) средств, которое осуществляют или с помощью маски...
Как мы говорим и как мы слушаем: общение можно сравнить с огромным зонтиком, под которым скрыто все...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Билет № 1
Метилирование в биохимических процессах. (Что используют метилазы(ТГФ-кислота, витамин В12), как происходит процесс, за счет чего, что метилируется)
Метилирование — введение в органические соединения метильной группы -СН3 вместо атома водорода, металла или галогена.
Витаминами B12 называют группу кобальтсодержащих биологически активных веществ, называемых кобаламинами. Витамин В12 участвует в двух видах реакций – реакции изомеризации и метилирования. Основой изомеризующего действия витамина В12 является возможность способствовать переносу атома водорода на атом углерода в обмен на какую-либо группу. Участвует в трансметилировании аминокислоты гомоцистеина при синтезе метионина. Метионин в дальнейшем активируется и используется для синтеза адреналина, креатина, холина, фосфатидилхолина и др.
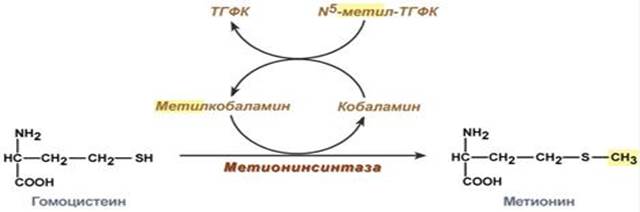
У эукариот метилирование ДНК – один из ключевых механизмов регуляции онтогенеза и клеточной дифференцировки, а также подавления экспрессии чужеродных последовательностей и мобильных элементов. После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей ДНК, при этом образуется N6-метиладенин, а также N5-метилцитозин.
Метилирование цитозина и аденина
• Не нарушает способность к комплементарному взаимодействию, но стабилизирует двойную спираль ДНК и распознается многочисленными белками
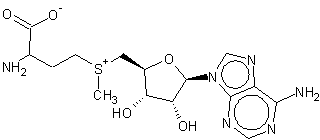
• Происходит с участием S-аденозилметионина (SAM) (донор метильной группы), может происходить “спонтанно” без участия ферментов.SAM
• В результате дезаминирования метилированного цитозина возникает тимин,
что часто происходит в клетке и приводит к мутации, закрепляемой при
|
|
репликации ДНК.
Наличие метильных групп в цепях ДНК необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов. Метилирование может быть использованно в процессе секвенирования ДНК.
Стерео изомерия моносахаридов. (Что это такое, что под этим понимают. Чем отличаются гексозы/пентозы, L/D изомеры, α/β)
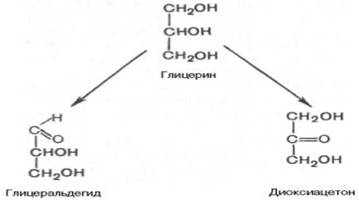
Моносахариды можно рассматривать как производные многоатомных спиртов, содержащие карбонильную (альдегидную или кетонную) группу. Если карбонильная группа находится в конце цепи, то моносахарид представляет собой альдегид и называется альдозой; при любом другом положении этой группы моносахарид является кетоном и называется кетозой. Простейшие представители моносахаридов – триозы: глицеральдегид и диоксиацетон. При окислении первичной спиртовой группы трехатомного спирта – глицерола – образуется глицеральдегид (альдоза), а окисление вторичной спиртовой группы приводит к образованию диоксиацетона (кетоза).
Стереоизомерия моносахаридов. Все моносахариды содержат асимметричные атомы углерода: (к-во асимметрич. атомов у альдоз: к-воС-2) альдотриозы – один центр асимметрии, альдо-тетрозы – 2, альдопентозы – 3, альдогексозы – 4 и т.д. Кетозы содержат на один асимметричный атом меньше(nC-3), чем альдозы с тем же числом углеродных атомов. Следовательно, кетотриоза диоксиацетон не содержит асимметричных атомов углерода. Все остальные моносахариды могут существовать в виде различных стереоизомеров. Общее число стереоизомеров для любого моносахарида выражается формулой N = 2n, где N – число стереоизомеров, а n – число асимметричных атомов углерода. Глицеральдегид содержит только один асимметричный атом углерода и поэтому может существовать в виде двух различных стереоизомеров. Изомер глицеральдегида, у которого при проекции модели на плоскость ОН-группа у асимметричного атома углерода расположена с правой стороны, принято считать D-глицеральдегидом, а зеркальное отражение – L-глицеральдегидом:
|
|

Наиболее типаичными для живых организмов являются D моносахариды. Альдогексозы содержат четыре асимметричных атома углерода и могут существовать в виде 16 стереоизомеров(24), представителем которых является, например, глюкоза. Для альдопентоз и альдотетроз число стереоизомеров равно соответственно 23 = 8 и 22 = 4.
Оптическая изомерия. Природные моносахариды обладают оптической активностью. Способность вращать плоскость поляризованного луча света – одна из важнейших особенностей моносахаридов, молекулы которых имеют асимметричный атом углерода или асимметричны в целом. Свойство вращать плоскость поляризованного луча вправо обозначают знаком плюс (+), а в противоположную сторону – знаком минус (–). Так, D-глицеральдегид вращает плоскость поляризованного луча вправо, т. е. D-глицеральдегид является D(+)-альдотриозой, а L-глицеральдегид – L(–)-альдотриозой. Моносахариды, относящиеся по стереохимической конфигурации к D-ряду, могут быть левовращающими(-).
Циклические (полуацетальные) формы моносахаридов. Шестичленные кольца сахаров называют пиранозами, а пятичленные – фуранозами. α-Форма – это форма, у которой расположение полуацетального гидроксила такое же, как у асимметричного углеродного атома, определяющего принадлежность к D- или L-ряду. Иными словами, в формулах с α-модификацией моносахаридов D-ряда полуацетальный гидроксил пишут снизу, а в формулах представителей L-ряда – сверху. При написании β-формы поступают наоборот. Таким образом, явление мутаротации связано с тем, что каждый твердый препарат углеводов представляет собой какую-либо одну циклическую (полуацетальную) форму, но при растворении и стоянии растворов эта форма через альдегидную превращается в другие таутомерные циклические формы до достижения состояния равновесия. Например, известно, что в водных растворах глюкоза находится главным образом в виде α- и β-глюкопираноз, в меньшей степени – в виде α- и β-глюкофураноз и совсем небольшое количество глюкозы – в виде альдегидной формы. Различные таутомерные формы глюкозы в свободном состоянии известны лишь α- и β-пиранозы. Существование малых количеств фураноз и альдегидной формы в растворах доказано, но в свободном состоянии они не могут быть выделены вследствие своей неустойчивости.
|
|
Изменение величины удельного вращения при стоянии (во времени) растворов моносахаридов называется мутаротацией(переход от α к β). Мутаротация должна вызываться изменением асимметрии молекулы, а следовательно, трансформацией ее структуры в растворе.

Билет № 2
Челночные механизмы переноса веществ через мембрану. (Главным образом речь о митохондриях. Перенос АТФ, АДФ, жирных кислот, пировиноградной кислоты через мембрану)
Образующийся в цитоплазме NADH не может самостоятельно проникать через внутреннюю митохондриальную мембрану (ВММ) в матрикс (внутреннее пространство митохондрии), где он должен утилизироваться в дыхательной цепи, поставляя в нее электроны. Эту функцию выполняет целый ряд челночных механизмов, из которых два заслуживают наибольшего внимания.
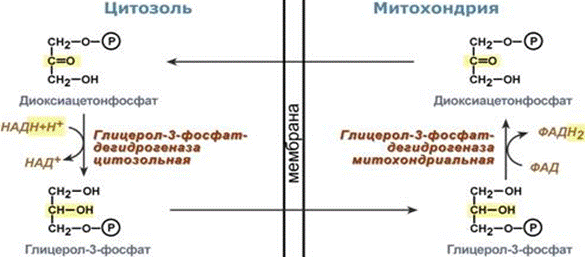
Глицерофосфатный челнок. Работает в белых скелетных мышцах, мозге, в жировой ткани, гепатоцитах. Дигидроксиацетон-3-фосфат взаимодействует с цитоплазматическим NADH с образованием глицерин-3-фосфата:
СН2(ОН)-С(=О)-СН2-ОРО3Н2 + NADH+H+ = СН2(ОН)-СH(ОH)-СН-ОРО3Н2 + NAD+
Глицерин-3-фосфат передает электроны на FMN(флавопротеин содержащий ФАД), локализованный во внутримитохондриальной мембране, по схеме:
FMN + СН2(ОН)-СH(ОH)-СН-ОРО3Н2 = СН2(ОН)-С(=О)-СН-ОРО3Н2 + FMNH2(передает 2 электрона вместе с 2Н+)
Глицерофосфатный механизм передает электроны ТОЛЬКО ВНУТРЬ митохондрий.
Аспартат-малатный челнок. является универсальным, работает в печени, почках, сердце
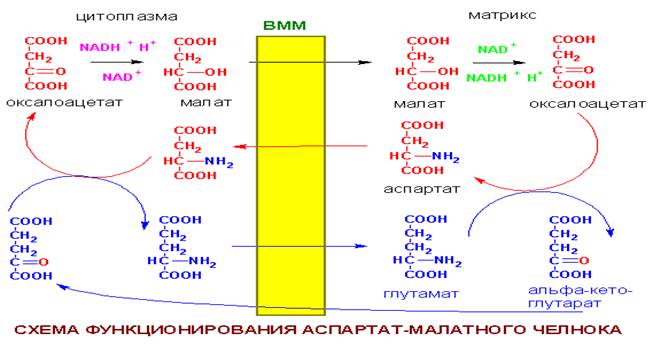
Из перечисленных соединений аспартат-малатного челнока только оксалоацетат и NADH не могут самостоятельно пересекать ВММ (не имеют встроенных в эту мембрану переносчиков). Для обеспечения функционирования челнока и требуются реакции переаминирования(перенос аминогруппы с одной молекулы на другую).
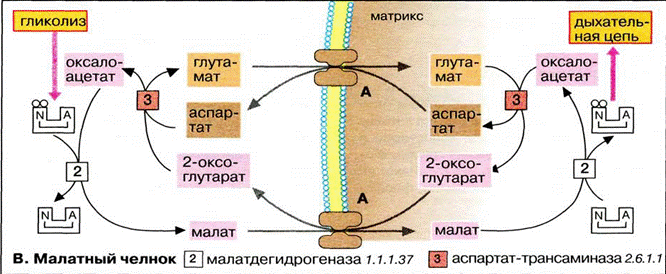
Разница между двумя механизмами переноса восстановительных эквивалентов заключается в том, что аспартат-малатный челнок поставляет в дыхательную цепь молекулы NADH, фигурирующие в начальных ее этапах. При переносе по дыхательной цепи пары электронов, полученных из молекулы NADH, синтезируется в итоге 3 молекулы АТФ. Глицерофосфатный челнок поставляет в цепь молекулы FADH2, в результате чего из одной цитоплазматической NADH синтезируется только 2 молекулы АТФ (FADH2 располагается на более низких "энергетических ступенях" дыхательной цепи).
|
|
В митохондриях за перенос жирных кислот отвечает специальная транспортная система. Активированные жирные кислоты в форме ацетил-КоА становятся транспортабельными в цитоплазме после взаимодействия с карнитином. Образовавшийся ацилкарнитин транспортируется в матриксе карнитиновым переносчиком, обмениваясь на свободный карнитин. В матриксе ацильные остатки вновь связываются с КоА.
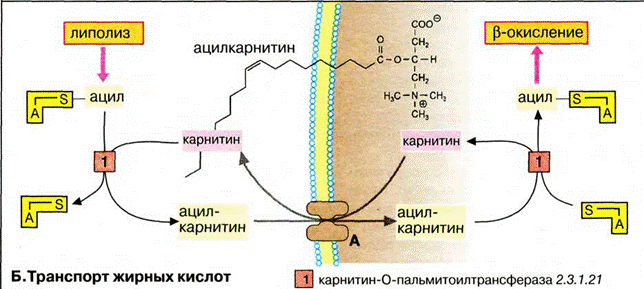
Окисление жирных кислот. (про α-окисление β-окисление)
При попадании в клетки жирные кислоты активируются путем образования ацетил-КоА. Для этого нужны две связи АТФ. В матрикс митохондрий активированные жирные кислотытранспортируются в виде ацилкарнитина, который является трансмембранным переносчиком. Деградация жирных кислот происходит в митохондриальном матриксе путем окислительного цикла реакций, при котором последовательно отщепляются С2-звенья в виде ацетил-КоА(активированной уксусной кислоты). Последовательное отщепление ацетильных групп начинается с карбоксильного конца активированных жирных кислот каждый раз между С-2(альфа-атомом) и С-3(β-атомом). Поэтому цикл и называется бета-окислением, пространственно и функционально бета-окисление связано с цитратным циклом и дыхательной цепью.
● Первая стадия - дегидрирование активированной жирной кислоты(ацил-КоА) с образованием β-ненасыщенной жирной кислоты с двойной связью в транс-конфигурации. При этом оба атома водорода с электронами переносятся от фермента(1) на электронопереносящий флавопротеин(ETF). ETF-дегидрогеназа(5) переносит восстановительные эквиваленты на убихинон(кофермент Q), который является составной частью дыхательной цепи.
● Вторая стадия деградации жирной кислоты состоит в присоединении молекулы воды к двойной связи ненасыщенной жирной кислоты(р-ция 2 - гидратирование)
● На третьей стадии происходит окисление гидроксильной группы при С-3 в карбонильную(р-ция 3 - дегидрирование). Акцептором восстановительных эквивалентов является НАД+, который передает их в дыхательную цепь.
● На четвертой стадии активированная β-кетокислота расщепляется ацетилтрансферазой в присутствии кофермента(р-ция 4 - тиолитическое расщепление). Продуктами реакции являются ацетил-КоаА и активированная жирная кислота, углеродная цепь которой короче на 2(два) углеродных атома по сравнению с длиной цепи исходной кислоты. Для полной деградации длинноцепочечной жирной кислоты цикл должен многократно повторяться; например для стеарил-КоА(18:0) - 8 раз.
|
|
● α-окисление - разрушаются метилразветвленные жирные кислоты. Процесс начинается с гидроксилирования и далее осуществляется путем последовательного отщепления С1-остатков, не требует участия кофермента А и не сопровождается синтезом АТФ.
● ω-окисление - начинается с гидроксилирования омега-углеродного атома и в результат приводит к образованию жирных кислот с двумя карбоксильными группами, которые разрушаются бета-окислением с обеих сторон до С8 или С6-дикарбоновых кислот и выводятся с мочой.

Билет № 3
Нуклеотидные коферменты. (Какие, НАД(никотинамидадениндинуклеотид), ФАД(флавинадениндинуклеотид))
Особую функции выполняют в клетке некоторые моно- и динуклеотиды — вещества, близкие по своей структуре к компонентам нуклеиновых кислот. В состав каждого из них входят: фосфорная кислота, рибоза и пуриновое или пиримидиновое основание. Каждое из этих оснований может образовать нуклеозидтрифосфат, в котором последовательно соединены основание, сахар и три фосфатные группы.
Аденозинтрифосфат (АТФ), состоящий из аденина, рибозы и трех остатков фосфата, играет важнейшую роль как своего рода «энергетическая валюта» всех клеток.
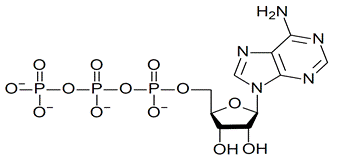
В молекуле две концевые фосфатные группы присоединены к нуклеотиду макроэргическими (т. е. богатыми энергией) связями, которые обозначаются символом — Ф. Биологически полезная энергия этих связей может передаваться другим молекулам; большая часть химической энергии клетки запасается в форме макроэргических фосфатных связей АТФ и может использоваться клеткой при переносе фосфатной группы на другую молекулу.
Гуанозинтрифосфат (ГТФ) необходим на некоторых этапах синтеза белков, уридинтрифосфат (УТФ) — на некоторых этапах углеводного обмена, а цитидинтрифосфат (ЦТФ) — для синтеза жиров, и фосфолипидов. Трифосфаты всех четырех нуклеотидов нужны для синтеза РНК, а четыре соответствующих нуклеозидтрифосфата, содержащие дезоксирибозу, — для синтеза ДНК.
 ГТФ
ГТФ
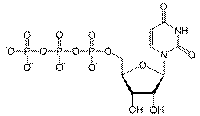 УТФ
УТФ 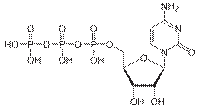 ЦТФ
ЦТФ
Перечень нуклеотидов, играющих важную роль в метаболических процессах, завершают НАД, НАДФ и ФАД.
Никотинамидадениндинуклеотид (НАД), называемый также дифосфопиридиннуклеотидом (ДПН), состоит из никотинамида, рибозы и фосфата, присоединенных к адениннуклеотиду (состоящему из фосфата, рибозы и аденина). НАД имеет первостепенное значение как первичный акцептор электронов и водорода в окислительных реакциях, протекающих внутри клетки. Ферменты дегидрогеназы отнимают электроны и водород от молекул таких веществ, как, например, молочная кислота, и переносят их на НАД, который в свою очередь передает их другим акцепторам электронов. Для некоторых других ферментов акцептором электронов и водорода служит никотинамидадениндинуклеотидфосфат (НАДФ), называемый также трифосфопиридиннуклеотидом (ТПН). Он отличается от НАД только тем, что имеет третью фосфатную группу, присоединенную к рибозе адениннуклеотида.
 НАД+
НАД+
Флавинадениндинуклеоти д (ФАД) (состоящий из соединенных в цепочку рибофлавина, рибита, двух фосфатных групп, рибозы и аденина) служит акцептором водорода и электронов для некоторых других дегидрогеназ. Следует особо отметить, что в состав этих динуклеотидов входят витамины — никотинамид и рибофлавин. ФАД
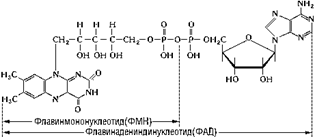
Такие вещества, как НАД, НАДФ и ФАД, называются коферментами; они необходимы для функционирования определенных ферментных систем, но очень непрочно связаны с молекулой фермента и легко отщепляются от нее. Присоединяя электроны и атомы водорода, они переходят из окисленной формы в восстановленную; например, окисленная форма НАД (НАД+) превращается в восстановленную НАД • Н. Передав электроны следующему акцептору в цепи дыхательных ферментов, кофермент снова переходит в окисленную форму.
Пируватдегидрогеназный комплекс. (про коферменты(5) и ферменты(3) в составе комплекса, нарисовать их)
Пируватдегидрогеназный комплекс (ПДК) — комплекс трех ферментов, который осуществляет окислительное декарбоксилирование пирувата. Продуктами окисления являются углекислый газ, ацетил-КоА,
Компоненты: 3 фермента, 3 кофактора (простетическая группа, связанная с апоферментом прочно ковалентно), 2 кофермента (простетическая группа, связанная с апоферментом непрочно нековалентно):
1. Е1 —фермент пируватдегидрогеназа декарбоксилирующая(гидрирование пирувата).
Кофактором является простетическая группа —активная форма определенного витамина
 В1 — тиаминпирофосфат.
В1 — тиаминпирофосфат.
2. Е2 — фермент дигидролипоилацетилтрансфераза(переносит ацетил).
Кофактором является простетической группой является витаминоподобное вещество — липоевая кислота, которая присоединив 2 атома водорода может превращаться в дигидролипоил.
 Липоевая кислота
Липоевая кислота
Коферментом является активная форма пантотеновой кислоты — НS-КоА(кофермент А), она принимает ацетильный остаток от липоевой кислоты.
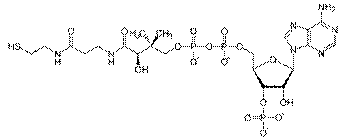 КоА
КоА
3. Е3 — дигидролипоилдегидрогеназа(отнимает Н).
Кофактором является простетическая группа флавинадениндинуклеотид (ФАД) — активная форма витамина В2.
 ФАД
ФАД
Коферментом является активная форма витамина PP — никотинамидадениндинуклеотид (НАД+).
 НАД
НАД
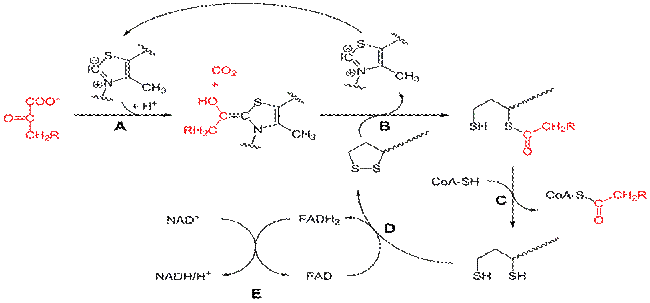
Работа пируватдегидрогеназного комплекса
Регуляция: «По механизму обратной связи» ингибируют работу пируватдегидрогеназного комплекса конечные продукты, увеличивает активность комплекса пируват.
Билет № 4
Билет № 5
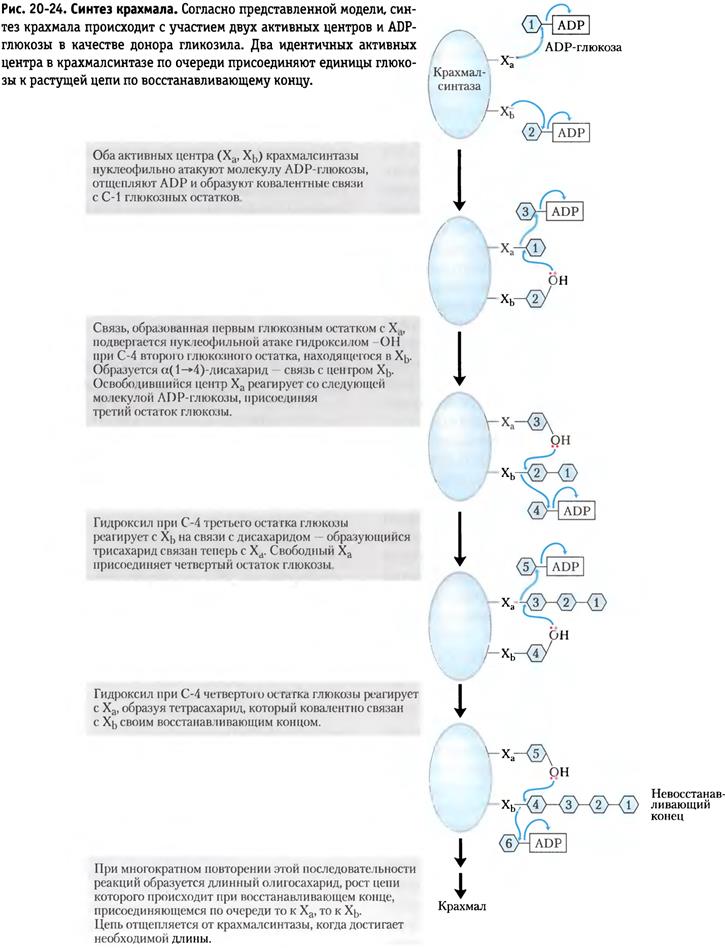
1. Роль нуклеотидов в биосинтезе полисахаридов. (Активизация сахаров, АТФ, U, G)
Общий механизм построения крахмала, где используются активированные АТФ молекулы глюкозы.
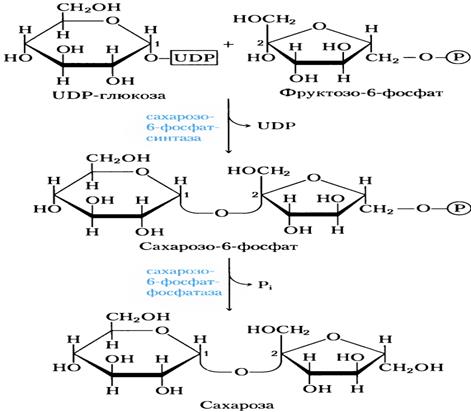
УТФ(урединтрифосфаот) участвует в активации глюкозы при синтезе дисахарида (сахарозы).
Для синтеза целлюлозы используется УДФ-глюкоза, синтез происходить внеклеточно в мембранном белке целлюлоз-синтазе.
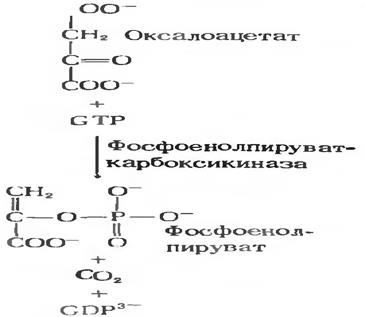
При глюконеогенезе ГТФ активирует ЩУК (оксалоацетат) и с помощью ФЕПкарбоксилазы получается ФЕП.
При синтезе гликогена, так же как и в синтезе целлюлозы используется УТФ, только гликоген в цитозоле.
Глюконеогене́з — метаболический путь, приводящий к образованию глюкозы из неуглеводных соединений (в частности, пирувата). Наряду с гликогенолизом, этот путь поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и органов.
Суммарное уравнение глюконеогенеза выглядит следующим образом:
2 Пируват + 4ATP + 2GTP + 2NADH + 2H+ + 4H2O → глюкоза + 4ADP + 2GDP + 6Pi + 2NAD+
Глиоксилатный цикл.
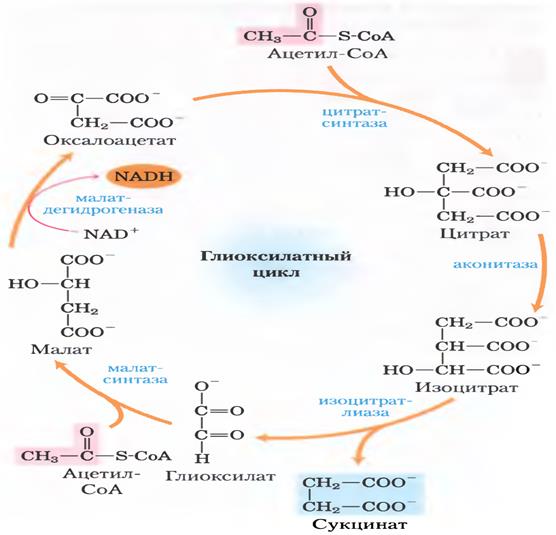

Сам цикл есть только у растений и некоторых беспов и конечно микроорганизмов. В растениях глиоксисомы в которых и происходит цикл, важно то что сукцинат может превращаться в оксалоацетат через фумарат и малат и оксалоацетат дальше в ФЕП(фосфоенолпируват), а ФЕП с помощью ФЕП-карбоксикиназы в глюкозу.
Билет 6
Билет 7
Классификация ферментов.

Оксидредуктазы: систематические названия их составляют по форме «донор: акцептор оксидоредуктаза» лактат-НАД оксидредуктаза отнимает электрон и протон у лактата, реакции ОВР
Трансферазы: наименование их составляется по форме «донор: транспортируемая группа – трансфераза» ацетилтрансферазы, амино-трансферазы, фосфотрансферазы.
Гидролазы: наименование их составляют по форме «субстрат-гидролаза»
Лиазы: «субстрат-лиазы»
Изомеразы: «субстрат – цис-транс-изомераза». Если изомеризация включает внутримолекулярный перенос группы, фермент получает название «мутаза»
Лигаза: название их составляют по форме «X:Y лигаза», где X и Y обозначают исходные вещества. Глутамат:аммиак лигазу сокращенное название «глутаминсинтетаза»
Билет № 8
Биосинтез пиримидиновых нуклеотидов. (Синтез двуцепочечный, сначала основание, потом прифигачивается сахар. Синтез в митохондриях(?))

РRPP-фосфорибозилпирофосфат
Сначала синтез пиримидинового кольца (Оротат), далее к нему присоединяется рибоза
Исходный УМФ фосфорилируется до ди-, а затем трифосфата УТФ. Он, в свою очередь, превращается цитидинтрифосфат-синтазой (СТР-синтаза) в ЦТФ.
Ферменты. Общая характерстика. (Характеристика фермента в цифрах: что такое Vм, Kм, скорость, активность. Регуляция)




Реакция Михаэлиса-Ментона
Лимитирующей по скорости реакцией является распад фермент-субстратного комплекса с образованием продукта
Математическая модель


При алгебраическом преобразовании математической модели 
Мы можем получить следующую кривую 
 под числом оборотов понимается,количество изменений конформации активного центра с образование продукта.
под числом оборотов понимается,количество изменений конформации активного центра с образование продукта.
Субстратная специфичность ферментов
Кратко про аллостерическое активация и ингибирование
Билет № 9
Билет 10.
Билет № 11
Билет № 12
Окислительное фосфорилирование. (Связано с дыхательной цепью. Транспорт протона, передача субстрату за счет соединения, дельта G которого повышается)
Окислительное фосфорилирование это энергозапасающий процесс являющийсяч финальной стадией клеточного дыхания. Протекает в митохондриях у эукариот. Суть процесса сводится к востановлению О2 до Н2О за счет переноса электронов от НАДН и ФАДН2. В процессе окисления глюкозы после гликолиза из одной молекулы получаются 2 молекулы пирувата идущие в митохондрии. Из молекул пирувата в пируватдегидрогеназном комплексе получается ацетил КоА. Он вступает в цикл Кребса в котором окисляется до СО2 и воды, что сопровождается получением АТФ и востановительных эквивалентов (НАДН и ФАДН2). Электрон полученный путем окисления восстановительного эквивалента поступает в электронно-транспортную цепь дыхания. Дыхательная цепь митохондрий состоит из ряда последовательно действующих переносчиков электронов, в том числе некоторых интегральных белков мембраны.
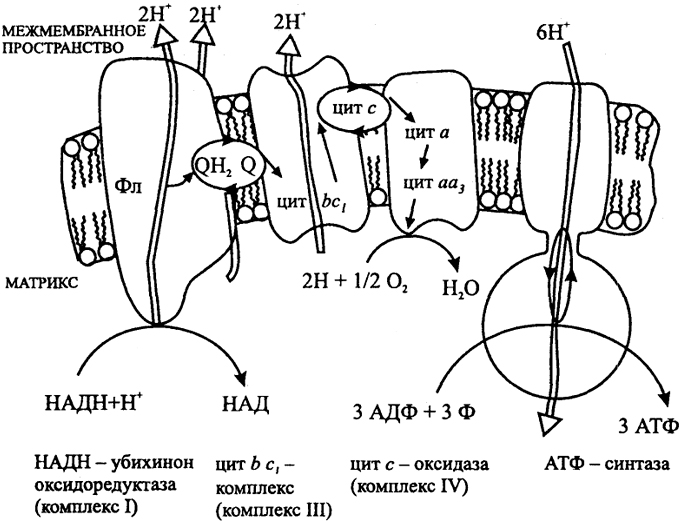

Суть цикла: электроны, полученные в результате окисления НАДН передаются на белок флавопротеин в составе комплекса НАДН убихинонредуктазы(комплекс 1). Далее электроны переносятся на железосерные белки, а с них на хинон вместе с Н+ из матрикса. К хинону также приходят электроны из комплекса 2, полученные путем окисления сукцината до фумарата. Хинон переносит электроны в Q-цикл в процессе которого электроны переносятся на цитохром С, а протоны в межмембранное пространство. Цитохром С переносит электроны в комплекс 4, который через гемы передает их кислороду(при этом из матрикса берутся недостающие протоны). Кислород востанавливается до воды. При этом выделяемая в процессе энергия используется для переноса протонов в межмембранное пространство. Итог- энергия высвобождаемая при переносе электронов в дых цепи запасается в виде протонного градиента.
Этот протонный градиент фоормирует протон движущую силу которая служит источником энергии для синтеза АТФ, когда ионы Н+ спонтанно возвразаются по граадиенту концентрации из цитозоля в матрикс, проходя через поры для Н+ в АТФ синтазе. АТФ синтаза состоиз F1(переферический мембранный белок) и F0(интегральный белок). Процесс синтеза АТФ проходит по механизму вращательного катализа: при каждом переносе Н+ в матрикс через F0 происходит циклическое изменение конформации 3-х центров связывания нуклеотидов в F1. Центры принимают конформацию благоприятную для связывания АДФ и фосфата, АТФ, пустой конформации, освобождающей нуклеотид. При этом энергия протон движущей силы расходуется на высвобождения АТФ из центра связывания. Активность окислительного фосфорилироввания зависит от соотнощения АТФ/АДФ.
Олигосахариды. Синтез. Значение. (Арабиноза. Связь инозита с галактозой. Синтез за счет активирования моносахаридов при связи с нуклеотидами)
Олигосахариды - это углеводы, состоящие из коротких цепей моносахаридных звеньев, соединенных гликозидными связями(связь, образуемая между OН группой одной молекулы и аномерным атомом(полуацетальным) другой молекулы с высвобождением молекулы воды).Наиболее распространенные дисахариды (сахароза:глюкоза+фруктоза). L-арабиноза????. Рафиноза (раффиноза) — невосстанавливающий трисахарид, состоящий из остатков D-галактозы, D-глюкозы и D-фруктозы., соединения между ними происходит через антомерные атомы
 Сахароза
Сахароза  Арабиноза
Арабиноза 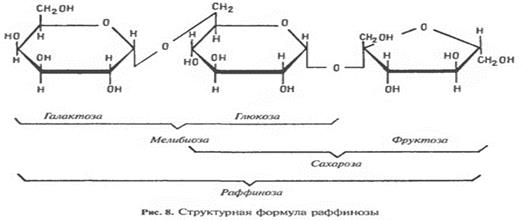
В живых организмах под действием аденозинтрифосфорной кислоты (АТФ) с участием ферментов происходит избирательное фосфорилирование только по гидроксильной группе при атоме С-6 глюкозы с образованием глюкозо-6-фосфата:
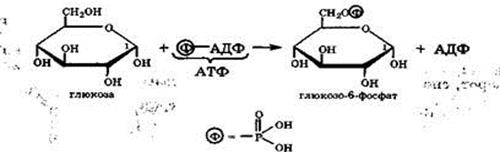
Значение: запасание питательных веществ (в вакуолях растений например), форма транспортировки питательных веществ. Сахароза является основным субстратом для синтеза целлюлозы, в мембранном комплексном белке (сахароз-синтаза и целлюлоз-синтаза) она разрушается и синтезируется целлюлоза.
Билет 13
Биосинтез жирных кислот
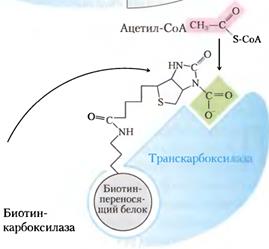
Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца.
HCO3- + АТФ +биотин= карбоксилированный биотин. Карбоксилированный биотин + CH3-CO-S-КоА= HOOC-CH2-CO-S-КoА. Катализирует эту реакцию фермент ацетил-КоА-карбоксилаза. Фермент содержит в качестве простетической группы биотин. Реакция протекает в два этапа: I – карбоксилирование биотина с участием АТФ и II – перенос карбоксильной группы на ацетил-КоА, в результате чего образуется малонил-КоА. Малонил-КоА представляет собой первый специфический продукт биосинтеза жирных кислот. В присутствии соответствующей ферментной системы малонил-КоА быстро превращается в жирные кислоты. Путем реакций малонилКоА и ацетил КоА с ацилпереносящим белком(АПБ) ацетил и малонил переносятся на АПБ, далее ацетилАПБ и малонилАПБ обединяются в ацетоацетилАПБ с потерей СО2. Цикл реакций повторяется вплоть до получения цепочки нужной длины.


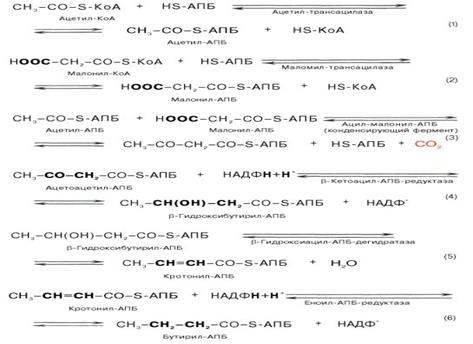


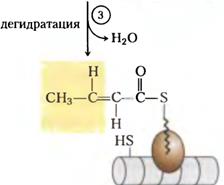

Видно при последовательном восстановлении можно оставлять двойные связи для синтеза полиненасыщенных жирных кислот
Запасные и структурные полисахариды растений Полисахариды -полимерные молекулы состоящие из большого количества остатков моносахаридов соединенных гликозидными связями(связь между аномерным атомом одной молекулы и ОН группы другой молекулы). Бывают гомополисахариды(один тип моносахаридов) и гетерополисахариды.
Функции: запасающая (крахмал, гликоген), структурная(целлюлоза, хитин)
Крахмал - смесь двух полимеров глюкозы: амилозы(длинные неразветвленные цепи из остатков D глюкозы соединенных альфа 1-4 связями) и амилопектина(длинные неразветвленные цепи из остатков D глюкозы соединенных как альфа 1-4, так и альфа 1-6 связями, через каждые 24-30 остатков разветвление). Тк связи в крахмале затрагивают один аномерный атом, следовательно краевые остатки глюкозы имеют свободные аномерные атомы, обладающие высокой хим. активностью.
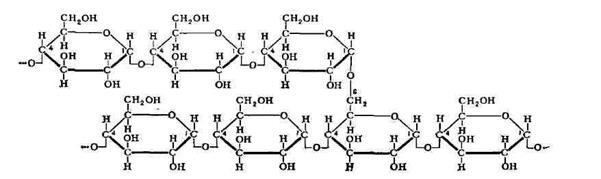 Крахмал
Крахмал
Целлюлоза - основной компонент клеточной стенок растений. Является линейным неразветвленным гомополисахаридом из 10-15 тыс. остатков D глюкозы, но связи между остатками бетта 1-4
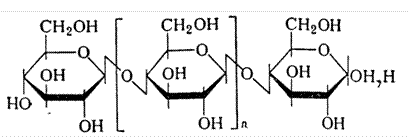 фрагмент целлюлозы
фрагмент целлюлозы
Каллоза - нерастворимый в воде полисахарид, содержащийся в растениях и состоящий из остатков молекулы глюкозы, соединённых в спиральную цепочку (в отличие от целлюлозы, в которой молекулы глюкозы соединены в прямую цепочку). Каллоза выстилает канальцы ситовидных пластинок флоэмы; по мере старения ситовидных трубок количество каллозы увеличивается, и ситовидные трубки закупориваются и перестают функционировать. При поранении каллоза откладывается на стенках клеток паренхимы, образуя каллюс. Встречается также в стенках клеток некоторых водорослей и грибов
Билет 14
Углеводы. Строение, свойства. Углеводы (сахара, сахариды) — органические вещества, содержащие карбонильную группу и несколько гидроксильных групп. Источником углеводов для всех живых организмов является процесс фотосинтеза, осуществляемый растениями. Углеводы — весьма обширный класс органических соединений, среди них встречаются вещества с сильно различающимися свойствами. Все углеводы состоят из отдельных «единиц», которыми являются сахариды. По способности к гидролизу на мономеры углеводы делятся на две группы: простые и сложные. Углеводы, содержащие одну единицу, называются моносахариды, две единицы – дисахариды, от двух до десяти единиц — олигосахариды, а более десяти — полисахариды.
Моносахари́ды — простейшие углеводы, не гидролизующиеся с образованием более простых углеводов — обычно представляют собой бесцветные, легко растворимые в воде, плохо — в спирте и совсем нерастворимые в эфире, твёрдые прозрачные органические соединения, одна из основных групп углеводов, самая простая форма сахара. Водные растворы имеют нейтральную pH. Некоторые моносахариды обладают сладким вкусом. Моносахариды содержат карбонильную (альдегидную или кетонную) группу, поэтому их можно рассматривать как производные многоатомных спиртов. Моносахарид, у которого карбонильная группа расположена в конце цепи, представляет собой альдегид и называется альдоза. При любом другом положении карбонильной группы моносахарид является кетоном и называется кетоза. В зависимости от длины углеродной цепи (от трёх до десяти атомов) различают триозы, тетрозы, пентозы, гексозы, гептозы и так далее.

Дисахари́ды — сложные органические соединения, одна из основных групп углеводов, при гидролизе каждая молекула распадается на две молекулы моносахаридов, являются частным случаем олигосахаридов. По строению дисахариды представляют собой гликозиды, в которых две молекулы моносахаридов соединены друг с другом гликозидной связью, образованной в результате взаимодействия гидроксильных групп (двух полуацетальных или одной полуацетальной и одной спиртовой). В зависимости от строения дисахариды делятся на две группы: восстанавливающие и невосстанавливающие. Например, в молекуле мальтозы у второго остатка моносахарида (глюкозы) имеется свободный полуацетальный гидроксил, придающий данному дисахариду восстанавливающие свойства. Дисахариды наряду с полисахаридами являются одним из основных источников углеводов в рационе человека и животных..
 Сахароза
Сахароза
Олигосахариды — углеводы, молекулы которых синтезированы из 2 — 10 остатков моносахаридов, соединённых гликозидными связями. Соответственно различают: дисахариды, трисахариды и так далее.
Наиболее распространены среди олигосахаридов дисахариды. Среди природных трисахаридов наиболее распространена рафиноза — невосстанавливающий олигосахарид, содержащий остатки фруктозы, глюкозы и галактозы — в больших количествах содержится в сахарной свёкле и во многих других растениях.

Полисахариды — общее название класса сложных высокомолекулярных углеводов, молекулы которых состоят из десятков, сотен или тысяч мономеров — моносахаридов. С точки зрения общих принципов строения в
группе полисахаридов возможно различить гомополисахариды, синтезированные из однотипных моносахаридных единиц и гетерополисахариды, для которых характерно наличие двух или нескольких типов мономерных остатков. Гомополисахариды (гликаны), состоящие из остатков одного моносахарида, могут быть гексозами или пентозами, то есть в качестве мономера может быть использована гексоза или пентоза. В зависимости от химической природы полисахарида различают глюканы (из остатков глюкозы), маннаны (из маннозы), галактаны (из галактозы) и другие подобные соединения. К группе гомополисахаридов относятся органические соединения растительного (крахмал, целлюлоза, пектиновые вещества), животного (гликоген, хитин) и бактериального (декстраны) происхождения.
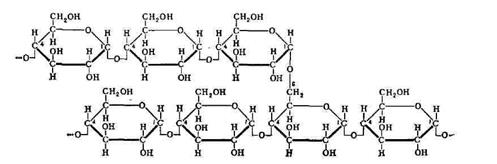 Крахмал
Крахмал
целлюлоза
Билет 15
Моносахариды. — простейшие углеводы, не гидролизующиеся с образованием более простых углеводов — обычно представляют собой бесцветные, легко растворимые в воде, плохо — в спирте и совсем нерастворимые в эфире, твёрдые прозрачные органические соединения, одна из основных групп углеводов, самая простая форма сахара. Водные растворы имеют нейтральную pH. Некоторые моносахариды обладают сладким вкусом. Моносахариды содержат карбонильную (альдегидную или кетонную) группу, поэтому их можно рассматривать как производные многоатомных спиртов. Моносахарид, у которого карбонильная группа расположена в конце цепи, представляет собой альдегид и называется альдоза. При любом другом положении карбонильной группы моносахарид является кетоном и называется кетоза. В зависимости от длины углеродной цепи (от трёх до десяти атомов) различают триозы, тетрозы, пентозы, гексозы, гептозы и так далее.


Здесь же показано и оптическая изомерия, заключающая в разности расположения заместителей при хиральном атоме.
Также возможна D и L изомерия определяется предпоследним атомом углерода и положением –ОН группы Д вправо Л влево
При сворачивании моносахаридов в циклы возможно образование альфа и бэта сахаров

Так при сворачивании моносахарида в цикл возможно образование как пиранозной (шестичленный цикл) так и фуранозной (пятичленный цикл) формы
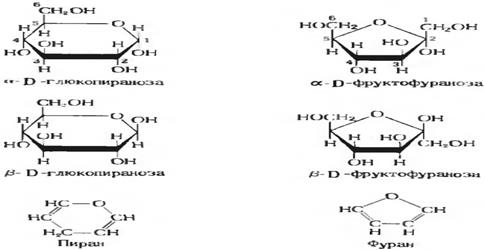
Моносахариды основной субстрат из которого получается энергия, можно сказать родоначальник биосинтеза, при метаболизме углеводов интермедиаты которого учавствуют во всех видах обмена (нуклеотидов, жиров, аминокислот и т.д.)
Краткие функии: энергетическая, кодификационная, структур
|
|
|

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой...

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...
Двойное оплодотворение у цветковых растений: Оплодотворение - это процесс слияния мужской и женской половых клеток с образованием зиготы...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!