Во всех прежних и даже современных учебниках по физиологии нет и намека на участие азота в дыхательной функции животных. Например, К. Шмидт-Ниельсен (1982) пишет «в большинстве физиологических процессов азот и инертные газы одинаково безразличны для организма». Однако И.М. Сеченов в статье «О газах крови» (1905) указывал, что содержание азота в крови составляет от 2 до 5%, т. е, он не только растворяется в плазме крови, но входит в химические соединения, участвующие в тканевом дыхании животных. Убедительные доказательства того, что в легких происходит не только обогащение эритроцитов кислородом и обеднение углекислотой, но и поглощение аминокислотами атмосферного азота, приводит М.И. Волский (1961). Вместе со своими сотрудниками он проводил эксперименты по инкубированию цыплят в таком воздухе, где азот был заменен аргоном; определял прирост азота в развивающемся курином эмбрионе и при развитии куколок пчел.
Оказалось, что яйца, заложенные в эксикатор, продуваемый аргонным воздухом, замирали на четвертый день, хотя при продувке обычным воздухом развивались нормально. Затем экспериментаторы перенесли нормальных цыплят сразу после вылупления в аргонный воздух. В обычном воздухе (контроль) цыплята чувствовали себя прекрасно, а в аргонном воздухе через шесть часов опускали крылья и через восемнадцать часов умирали. Поэтому ученые сделали заключение, что азот воздуха является не просто растворителем кислорода и нейтральным газом, а физиологически и биохимически активным веществом. Удалось также установить, что за счет азота воздуха во время инкубации цыпленка происходит прирост белка, который для яйца средней массы 60 г составляет 36 мг или 0,66%. Кроме того, была определена зависимость содержания азота от исходной массы яйца по контрольным и инкубированным яйцам. Аналогичные результаты были получены за период инкубации для куколок пчел, которые, не получая пищи, наращивают в своем организме азот. Таким образом, М.И. Волский считает, что настало время пересмотреть высказанное А. Лавуазье около 200 лет тому назад утверждение о том, что живые организмы не могут усваивать азот воздуха. По мнению автора, азот воздуха так же необходим животным, как и кислород, и можно полагать, что молекула гемоглобина наряду с кислородом захватывает газообразный азот, который идет на образование аминокислот, необходимых при синтезе белка
АДАПТАЦИОННЫЕ ОСОБЕННОСТИ ДЫХАНИЯ В РАЗЛИЧНЫХ УСЛОВИЯХ
Рыбы
Как говорилось выше, основной орган дыхания у рыб — жабры, они снабжают организм кислородом, передавая его из внешней водной среды в кровь. Однако у многих рыб водное дыхание спорадически дополняется воздушным, которое осуществляется в разной степени и через различные органы. Это явление следует рассматривать как приспособление для существования в среде с недостатком кислорода, т. е. в пресной воде при повышенных температурах.
Самая простая форма приспособления к использованию кислорода воздуха состоит в заглатывании воздуха ртом.. При понижении Ро2 в воде многие рыбы поднимаются к поверхности водоема и ртом заглатывают воздух вместе с верхним слоем воды, Вода в ротовой полости перемешивается с воздухом, насыщается кислородом, затем проходит через жабры. В этом случае не происходит формирования новых специальных органов дыхания, а жабры омываются водой, более обогащенной кислородом, чем вода толщи водоема. Значительную группу составляют рыбы, у которых временами при больших нехватках кислорода заглатываемый воздух обогащает кислородом кровеносную систему полости рта. При этом наблюдаются увеличение ротовой полости за счет специальных углублений и развитие сети капилляров.
У некоторых рыб возникло кишечное дыхание, при котором воздух, заглатываемый ртом, прогоняется через весь кишечник, на определенных участках которого имеется сильно развитая сеть кровеносных капилляров. Функция пищеварения и всасывания на таких участках не осуществляется, и они служат только для газообмена. У одних рыб отработанный воздух выходит через анус, у других через ротовую полость и затем через жаберные щели. Наиболее распространен (вьюновые, сомовые) тип кишечного дыхания, при котором газообмен происходит в средней и задней частях кишечника.
Соотношение вкладов основного (жаберного) и дополнительного (кишечного) дыхания меняется у разных рыб. Например, у вьюна доля кишечного дыхания почти равна доле жаберного, и поэтому кишечное дыхание необходимо, даже когда вьюн находится в хорошо аэрированной воде. Время от времени он поднимается на поверхность и заглатывает воздух, при этом ритм дыхания не учащается, а более интенсивно используется кишечное дыхание.
В процессе эволюции кишечное дыхание претерпело значительные изменения. Во-первых, ни у кого из позвоночных, кроме рыб, нет кишечного дыхания; во-вторых, значительное развитие получили специальные образования кишечной трубки, ее дериваты, такие, например, как плавательный пузырь. По своему происхождению плавательный пузырь может рассматриваться как простое выпячивание кишечной трубки с последующей специализацией. Она сводится к потере способности пищеварения и приобретению новых качеств: увеличения поверхности, газовой секреции и абсорбции кислорода из пузыря. Все эти приспособления служат для воздушного дыхания.
Всех рыб можно разделить на две большие группы: закрытопузырные, у которых плавательный пузырь не сообщается с кишечной трубкой, и открытопузырные, у которых связь пузыря с трубкой осуществляется посредством пневматического протока. Последние в течение всей жизни могут заглатывать воздух, который по пневматическому протоку проходит в плавательный пузырь. У личинок закрытопузырных рыб плавательный пузырь также сообщается с кишечной трубкой, и первое время рыбка заглатывает воздух, но вскоре проток атрофируется и плавательный пузырь становится закрытым. В дальнейшем у закрытопузырных рыб регуляция состава газов в пузыре происходит вследствие их секреции, а у открытопузырных главным образом в результате повторных заглатываний воздуха.
В случае необходимости плавательный пузырь может освободиться от газов. У открытопузырных освобождение происходит относительно просто. Газ из плавательного пузыря выдавливается через пневматический проток в пищевод, глотку и далее в окружающую воду, Использование кислорода, находящегося в плавательном пузыре, осуществляется через кровеносные сосуды и капилляры, проходящие по внутренней стенке пузыря.
В плавательном пузыре у закрытопузырных рыб имеются два образования, функция которых состоит в наполнении (секреции газов) и поглощении (резорбции газов). Первую функцию выполняет так называемое красное тело, или газовая железа, вторую — овал. Красное тело находится в переднем конце плавательного пузыря, а овал в заднем. Кровеносные сосуды, проходящие по внутренней стенке плавательного пузыря, в красном теле сильно ветвятся на ряд мелких, образуя густую сеть капилляров, что придает всему образованию красный цвет. Таким образом, красное тело выделяет в плавательный пузырь газ, который доходит до овала, где и происходит его резорбция. На дне овала проходят многочисленные сосуды и капилляры, и его можно рассматривать как второй сосудистый центр плавательного пузыря.
В отличие от других участков плавательного пузыря овал имеет кольцевую и радиальную группы мышечных волокон, расположенных так, что они создают сфинктер овала, Сфинктер может сокращаться и расслабляться, вследствие чего дно овала то закрывается, то открывается. Когда овал открыт, то газы легко могут резорбироваться сосудистой системой и поступать в общую систему кровообращения. Обычно плавательный пузырь используется для дыхания параллельно с жабрами. Например, у ильной рыбы, живущей в застойных водоемах Северной Америки, внутренняя поверхность пузыря увеличена за счет ячеистого строения. При низкой температуре воды (10°С) эта рыба обходится водным дыханием через жабры, а в нагретой до 30° С воде она совершает подъемы на поверхность каждые одну-две минуты и получает через плавательный пузырь 2/3 потребляемого кислорода, хотя углекислый газ выводится в основном жабрами.
Сходным образом ведет себя панцирная щука — реликтовый представитель древней группы костистых рыб, которая встречается только в Северной Америке. В холодной воде, особенно зимой, когда озера на несколько месяцев покрываются льдом, эта рыба использует жаберный тип дыхания, а при температуре 20-25°С у нее преобладает воздушное дыхание с помощью плавательного пузыря, покрывающее примерно 70-80% всей потребности кислорода. Летом через пузырь выводится 8-10% углекислого газа, тогда как основная масса углекислого газа выводится через жабры.
У основания глотки двоякодышащих рыб располагаются одно или два легких; открывающихся в пищевод, которые гомологичны легким наземных позвоночных. Стенки легких складчатые и выстланы дыхательным эпителием. Расстояние между поверхностью эпителия и протекающей по легочным капиллярам кровью составляет всего 0,5 мкм. Однолегочные рыбы, например австралийский рогозуб (Leoceratodus), постоянно живут в воде и дышат жабрами. Лишь при ухудшении аэрации водоема или его пересыхании они показываются на суше, где вентилируют свое легкое воздухом. Двулегочные рыбы, представители семейства чешуйчатниковых не могут обходиться без воздушного дыхания.
У некоторых рыб для воздушного дыхания приспособлены глотка и жаберная полость. Например, у бычков дыхание во влажном воздухе обеспечивается кожей головы, ротовой и жаберной полостями. Слизистая оболочка этих полостей обильно снабжена кровеносными сосудами. Воздух забирается ртом, кислород поглощается в ротовой или жаберной полостях, а оставшиеся компоненты воздуха выбрасываются через рот. Это наиболее простое приспособление для воздушного дыхания.
Более сложное приспособление представляет собой наджаберный орган, который обнаружен у змееголова, обитающего в Амуре. Этот орган образован выпячиванием глотки, а не собственно жаберной полостью. Наджаберный орган расположен над истинно жаберной полостью и соединяется с ней перед первой жаберной дугой. Наджаберная полость обильно выстлана слизистой оболочкой, снабженной сетью кровеносных капилляров. Воздух забирается ртом, и кислород поглощается сетью кровеносных сосудов и капилляров. В очень жаркие дни змееголов выплывает греться на обмелевшие берега к траве, а иногда ложится на водную растительность. Он долго лежит спокойно, причем движения жаберных крышек незаметны. В этих условиях воздушное дыхание у змееголова или полностью замещает жаберное или является весьма существенным дополнением к нему. Для нормальной жизнедеятельности змееголову необходим как атмосферный кислород, так и кислород, растворенный в воде. Иными словами, ни водный (жабры), ни воздушный (наджаберный орган) тип дыхания в отдельности не может полностью обеспечить змееголова кислородом. Это возможно только при функционировании и того и другого органа.
У рыб теплых вод имеется приспособление для воздушного дыхания в виде лабиринта. Этот орган образован выпячиванием собственно жаберной полости, и к его поверхности подходит много кровеносных сосудов и капилляров. Воздух, захватываемый рыбой, попадает из полости рта в лабиринт и отдает там кислород в кровь. Лабиринт есть у многих рыб: анабаса, гурами, макроподов, бойцовых рыб, трихогастера, поликантиса и др. Рыбы, имеющие лабиринт для воздушного дыхания, так приспособились к использованию кислорода воздуха, что даже в хорошо аэрируемой воде при нормальных условиях пользуются воздушным дыханием. Наиболее известна ползающая рыба (окунь-ползун), которая лазает по деревьям. Анабас может в течение нескольких дней находиться вне воды во влажном воздухе, хотя его лабиринтовый орган относительно невелик. Крупный лабиринт у бойцовой рыбки: он тянется к хвосту, за заднюю часть головы и налегает на передний конец плавательного пузыря (рис. 8).
| |
| |  |
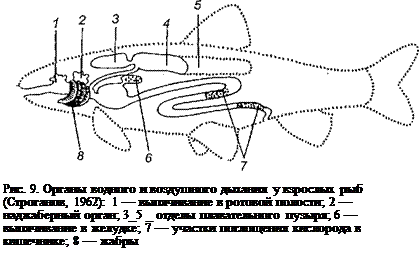
Такое обилие органов для водного и воздушного дыхания следует рассматривать как приспособление к жизни при недостатке кислорода в пресной воде при повышенных температурах. Наиболее удобными для расположения дыхательных поверхностей у рыб оказались ротовая полость и пищеварительный тракт, У некоторых рыб (вьюновые) средний и задний отделы кишечника стали участками резорбции кислорода, у большинства же рыб все органы дыхания находятся в задней части головы и передней части туловища (рис. 9), они тесно связаны с ротовой полостью и кишечной трубкой.
4. 2. Вторичноводные животные
Эволюционный процесс перехода в водную среду обитания у сухопутных животных начался очень давно. Предки наземных позвоночных животных ответвились в палеозое от кистеперых рыб, вышли на сушу и через стадию земноводных дали начало большому классу сухопутных животных — пресмыкающимся, от которых позже произошли классы теплокровных — птицы и млекопитающие. В каждом из классов наземных животных формировались и группы вторичноводных животных. Среди рептилий такими оказались мезозойские живородящие пресмыкающиеся — рыбоядные ихтиозавры, длина которых достигала 12 м, и прибрежные хищники плезиозавры длиной до 15 м. Первые походили на дельфинов, а вторые напоминали черепах с продетой через туловище змеей. Из ныне живущих рептилий к водным обитателям относятся морские и пресноводные черепахи, крокодилы и морские змеи; все они выходят на берег откладывать яйца, кроме некоторых живородящих морских змей,
Среди птиц группу вторичноводных ныряющих составляют утки, нырки, чистики, глупыши, олуши, буревестники, альбатросы, пингвины, гагары, кайры, бакланы, пеликаны и некоторые другие. Рекордсменами по погружению по праву считаются пингвины, которые ныряют на глубину до 125м.
Приспособления к нырянию в той или иной степени встречаются почти во всех основных отрядах млекопитающих, за исключением рукокрылых, зайцеобразных и приматов. Степень приспособления к жизни в водной среде у млекопитающих разных групп весьма различна и зависит от того, сколько времени они проводят в воде. Одни, поселившись на берегах водоемов, только на короткое время заходят в воду кормиться, чтобы спастись от врагов или найти прибежище в жаркие дни (например, утконос проводит в воде не более двух часов в сутки). Это лишь первые шаги освоения гидросферы частично водными животными, к которым принадлежат бобры, нутрии, ондатры, речные выдры и многие другие. Их обычно называют околоводными животными.
Более совершенны адаптации к погружению у полуводных млекопитающих — ластоногих и одного представителя хищных, калана. Они проводят большую часть жизни в воде, но еще тесно связаны с сущей, где рождают и выкармливают молоком детенышей, а также линяют и отдыхают. Хотя ластоногих обычно относят к полуводным млекопитающим, их уровень морфофизиологических адаптации к водной среде столь высок и так резко отличается от уровня околоводных животных, что представителей этой группы с полным правом можно считать водными млекопитающими Адаптация тюленей к водной среде настолько совершенна, что они могут даже спать под водой. Во время сна подъем на поверхность за новой порцией воздуха осуществляется рефлекторно.
Наконец, есть и исключительно водные млекопитающие — гидробионты, которые навсегда покинули берег, окончательно порвали с сушей и превратились в постоянных обитателей гидросферы. Это китообразные (киты и дельфины) и сиреновые (дюгони и ламантины). Китообразные наиболее сильно изменились в связи с водным образом жизни: приобрели рыбообразную форму тела и тем самым более всего уклонились от внешних форм наземных видов. По уровню специализации к жизни в водной среде и изменению внешних форм (веретенообразное тело, ластообразные передние конечности, горизонтальный хвостовой плавник, редуцированные задние конечности и т. д.) сиреновые больше приближаются к китообразным. У сирен и китообразных (неродственных групп), ведущих водный образ жизни с эоцена, возникло конвергентное сходство отдельных гомологичных органов. Однако оно более грубое, чем сходство признаков ряда близкородственных дивергентных форм.
Основной отличительной чертой поведения всех водных и полуводных млекопитающих является развитая в той или иной мере способность к нырянию. В зависимости от уровня специализации продолжительность ныряния и глубина погружения у них существенно различаются. Наиболее специализированные ныряющие млекопитающие — это многие китообразные и ластоногие. У них большая продолжительность ныряния, они способны при погружении достигать весьма значительных глубин. Рекордсмен по длительности погружения — бутылконос может пребывать под водой до 120 мин. Немногим ему уступает кашалот — 75 мин, но зато ему принадлежит рекорд по глубине погружения, по-видимому, до 2000 м. У мелких китообразных (дельфинов) в большинстве случаев продолжительность ныряния гораздо меньше, чем у крупных китов. Продолжительность ныряния дельфинов отдельных видов колеблется от 4 до 15 мин, а максимальная доступная им глубина обычно не превышает 100 м, иногда достигает 250-300 м. По продолжительности ныряния сиреновые значительно уступают многим китообразным, например, ламантин может пробыть под водой не дольше 16 мин. Глубинных погружений сиреновые обычно не совершают, так как питаются водной растительностью в прибрежной зоне.
Среди ластоногих имеются превосходные ныряльщики, сравнимые по продолжительности погружения со многими китообразными. Один из лучших ныряльщиков, тюлень Уэдделла может погружаться на глубину до 600 м, а кольчатая нерпа была поймана в ловушку на глубине 900 м. Максимальное время пребывания под водой колеблется у разных видов ластоногих от 20 до 45 мин.
Если сравнивать максимально возможные сроки пребывания под водой ныряющих млекопитающих и представителей наземных видов, то у последних это время заметно ниже. Например, нетренированный человек может пробыть под водой не более 1 мин. В то же время ловцы жемчуга и собиратели губок могут погружаться под воду на срок до 4 мин.
В чем же заключаются секреты продолжительного ныряния водных животных? При погружении под воду у них прекращается легочное дыхание, что должно было бы приводить к нарушению энергетического метаболизма. Однако этого не происходит, поскольку в процессе эволюции сформировался комплекс приспособлений, позволяющий вторичноводным животным переносить пребывание под водой без ущерба для жизнедеятельности.
По мнению В.П. Галанцева (1977, 1988), эволюционные адаптации затрагивают главным образом дыхательную и сердечнососудистую системы. У вторичноводных млекопитающих произошли значительные изменения в строении дыхательной системы, что выражается в уменьшении длины дыхательных путей, надежном разделении дыхательных и пищеварительных путей вследствие изменений в строении гортани и глотки, увеличении дыхательной поверхности легких, более обильной васкуляризации легочной ткани, установлении аритмичного дыхания с его длительной задержкой и ряде других приспособлений.
У всех китообразных и ластоногих, в отличие от наземных видов, отмечается очень энергичный и кратковременный дыхательный акт — выдох-вдох. Продолжительность этого акта у многих китообразных составляет 0,75-1 с. Объем вдыхаемого и выдыхаемого воздуха — дыхательный объем, по отношению к массе тела у многих ныряющих млекопитающих значительно больше, чем у человека и ряда наземных животных. Легкие высокоспециализированных ныряльщиков почти полностью опустошаются при выдохе, тогда как у человека при выдохе в легких остается 1/3 использованного воздуха (мертвое пространство). Например, у афалины за одно дыхание обновляется 90% всего легочного воздуха.
Для ныряющих млекопитающих типична высокая степень утилизации кислорода вдыхаемого воздуха. Это обеспечивается целым рядом особенностей: большой диффузионной поверхностью легких; длительными дыхательными паузами; высоким парциальным давлением кислорода в альвеолах (106-116 мм рт. ст.), большим объемом циркулирующей крови и др.
Содержание углекислого газа в выдыхаемом воздухе у крупных ныряющих животных обычно вдвое выше, чем у человека, а кислорода значительно ниже. За один дыхательный акт количество извлеченного из воздуха кислорода у дельфинов в несколько раз больше, чем у наземных животных. Кроме того, дыхательный центр вторичноводных животных менее чувствителен к накоплению углекислого газа в организме.
Глубоководные ныряльщики обычно уходят под воду на выдохе, к тому же объем их легких относительно невелик, что предохраняет животных от развития симптомов кессонной болезни. В то же время полуводные насекомоядные, грызуны, хищные, гиппопотамы и некоторые ластоногие, которые ныряют на небольшую глубину и на короткое время, обычно погружаются в воду на вдохе, так как кессонная болезнь им не грозит. Существенное значение в регуляции дыхания у них приобретают мышечные сфинктеры на бронхиолах и альвеолах. Сфинктеры препятствуют выходу воздуха в полость крупных бронхов, обеспечивая тем самым нормальное протекание газообменных процессов в респираторных отделах дыхательной системы.
Как же происходит обеспечение организма кислородом во время ныряния? Это связано с адаптационными перестройками в кровеносной и сосудистой системах. Прежде всего, следует отметить, что объем крови у ныряющих животных существенно больше, чем у наземных. Например, у тюленя массой 30 кг столько же крови, сколько у человека массой 70 кг. Больший объем крови, безусловно, играет важную роль в ее дыхательной функции, так как обеспечивает увеличение числа эритроцитов без чрезмерного сгущения крови, а значит, без нарушений ее микроциркуляции.
Кроме того, кровь у ныряющих животных отличается повышенной кислородной емкостью. Так, у дельфинов она на 25-30% выше, чем у наземных животных, и если у человека значение показателя составляет 20 мл кислорода на 100 мл крови, то у тюленей и китов 30-40 мл кислорода на 100 мл крови. Объясняется это большей концентрацией гемоглобина в крови водных и полуводных животных по сравнению с наземными млекопитающими. Например, у афалины в одном эритроците содержится 43,6% гемоглобина, что почти вдвое выше, чем у наземных.. Поэтому общая обеспеченность организма гемоглобином у ныряющих животных в расчете на единицу массы очень высока, почти вдвое выше, чем у наземных животных и составляет у некоторых тюленей 22-24 г/кг или 52 мл на 1 кг массы против 31 мл у человека. Кроме того, сродство гемоглобина к кислороду, определяемое степенью его насыщения при постоянном напряжении кислорода, у многих ныряющих животных выше, чем у наземных форм.
У большинства ныряющих животных отмечается высокое содержание в мышцах миоглобина, поэтому обычно их мышцы темные, например у китов его в 3-7 раз больше, чем у наземных. В мышцах тюленя содержание кислорода доходит до 47% общего количества кислорода в организме, доступного для метаболизма. Предполагалось, что ныряющим животным возможность долгое время находиться под водой обеспечивает кислород оксигемоглобина. Впоследствии, однако, было установлено, что миоглобин очень легко соединяется с кислородом, а его сродство к кислороду в 10 раз выше, чем гемоглобина крови. Поэтому при нормальной циркуляции крови в мышцах кислород из оксигемоглобина переходит в миоглобин, а не наоборот. Все это свидетельствует о том, что под водой высокое содержание кислорода в миоглобине у ныряющих животных может идти на покрытие потребности в кислороде исключительно лишь мышечной системы, которая затем переводится на анаэробный тип обмена.
В самых общих чертах можно сказать, что в мышечных клетках в процессе их работы идет гликолиз, в ходе которого глюкоза превращается в молочную кислоту. Такой анаэробный процесс способен дать значительное количество энергии для деятельности мышц, и именно в этом заключается одно из важных приспособлений, водных млекопитающих к длительному перерыву дыхания. Откуда же у китов берется огромное количество углеводов, необходимых для получения энергии в мышцах? Трудно представить, чтобы они все были заранее запасены в печени, с которой связаны огромные венозные коллекторы. Очевидно, немалое значение имеет то обстоятельство, что киты при погружении интенсивно питаются и переваривание пищи на глубине идет достаточно быстро, т. е. кровь немедленно насыщается углеводами.
Таким образом, хотя запасы кислорода в организме многих ныряльщиков почти вдвое больше, чем у человека, этого недостаточно для объяснения их способности задерживаться под водой гораздо дольше, чем может человек. Например, если общие запасы кислорода у тюленя массой 30 кг составляют 1520 мл, то за 25 мин в состоянии покоя он потребляет в четыре-пять раз больше кислорода, чем использует для ныряния; Следовательно, можно прийти к выводу, что при нырянии интенсивность потребления кислорода организмом существенно понижается в результате действия каких-то других механизмов. Решающая роль в снабжении и распределении кислорода в организме принадлежит при этом сердечно-сосудистой системе.
У всех ныряющих животных резко замедляется сердечная деятельность при погружении. Частота сердцебиений снижается у различных видов в 4-10 раз, а в отдельных случаях в 20 раз и более. Этот эффект, известный уже более 100 лет, получил название рефлекторной брадикардии. К настоящему времени брадикардия при нырянии описана как у высокоспециализированных ныряющих животных — тюленей, китов, дельфинов многих видов, так и у околоводных — утконоса., куторы, выхухоли, водяных полевок, ондатры, нутрии, бобра, норок, выдры и др.
Следует отметить, что интенсивность развития брадикардии при нырянии, видимо, не связана с уровнем адаптации животных и их способностью к длительному нахождению под водой. Так, наиболее резкое замедление сердечной деятельности при погружении свойственно нутрии (иногда в 20 раз и более), далеко не первоклассному ныряльщику. У норок и водяных полевок частота сердцебиений под водой замедляется в 4-5 раз, у ондатры в 5-10 раз, а у обыкновенного тюленя в 25 раз, со 100 до 4 ударов в 1 мин.
Наиболее слабо выраженная брадикардия обнаруживается при нырянии некоторых дельфинов. Так, по данным В.П. Галанцева, у дельфинов-азовок частота пульса при погружении снижается на 42%, а у дельфина-афалины на 50% (рис. 10). У некоторых других китообразных частота сердцебиений уменьшается в пять раз.
У всех ныряющих животных под водой частота сердечного ритма отличается изменчивостью и нерегулярностью, т. е. наблюдается аритмия на фоне выраженной брадикардии. Снижение частоты пульса при нырянии связано с изменением тонуса блуждающих нервов. Замедление пульсаций сердца, видимо, само по себе снижает его потребности в кислороде при нырянии. Кроме того, оно сопровождается ограничением или полным выключением кровоснабжения мышечной системы и многих других органов. Кровоток мало изменяется лишь в сосудах, снабжающих кровью головной мозг и сердечную мышцу. В результате перераспределения кровотока нервная система и сердечная мышца при нырянии обеспечиваются кислородом крови на том же уровне, что и до ныряния. Следует также отметить, что при нырянии происходит снижение интенсивности обмена веществ. Например, почки, которые в норме обильно снабжаются кровью и потребляют много кислорода, во время ныряния практически лишаются притока крови, и в них сразу прекращается клубочковая фильтрация и образование мочи. Сложному перераспределению кровотока в организме ныряющих животных способствуют морфологические изменения в кровеносной сосудистой системе. Особенно большие изменения произошли в венозной системе большого круга кровообращения. Перестройки сердечнососудистой системы ныряющих животных связаны с функциональными адаптациями и направлены в основном на обеспечение бесперебойного кровоснабжения головного мозга, депонирование крови в период развития брадикардии и перераспределение кровотока при погружении. Все изменения сосудистого русла шли, прежде всего, по линии образования венозных коллекторов и сплетений и развития особых путей оттока и притока крови к головному мозгу.
Вены, благодаря своим размерам и растянутости стенок, выступают идеальным регулирующим отделом всей системы кровообращения. Учитывая большой объем венозного русла и наличие венозных расширений (коллекторов) у ныряющих животных, можно заключить, что в них скапливается кровь.
Выше указывалось, что при нырянии происходит резкое снижение частоты сердечных сокращений и прекращение кровотока в периферических сосудах мышц и многих органов. В это время в венозных коллекторах начинает постепенно накапливаться кровь. Сужение просвета сосудов, подходящих к сердцу (в системе передних и задних полых вен), наличие диафрагмального сфинктера и особенности развития непарных вен дают основание говорить о регулирующем значении этих образований в поступлении крови к правому предсердию из заполненных кровью магистралей и коллекторов.
Для всех околоводных млекопитающих характерно увеличение калибра задней полой вены и ее ветвей, которые по диаметру во много раз превосходят соответствующие артерии. У ряда водных видов венозные ветви системы задней полой вены большей частью двойные и связаны между собой обширными анастомозами, что также увеличивает емкость венозного русла, выполняющего функцию депо крови на период ныряния.
У ластоногих основное депо крови — печеночный синус. Наполнение коллекторов при нырянии обеспечивается, в частности, рефлекторной брадикардией и ограничением кровотока в мышцах и многих органах. После выныривания возникающая тахикардия содействует опорожнению крови из коллекторов. Этому способствуют также возрастание разности артериального и венозного давления, увеличение минутного объема сердца и другие гемодинамические сдвиги.
К приспособительным особенностям сосудистого русла китообразных можно отнести венозные сплетения, развитые анастомозы между отдельными ветвями и специфические сосудистые образования в различных участках тела — чудесную сеть. Крупные сосуды местами разделяются на множество густо переплетенных между собой ветвей, которые образуют плотную сеть. Эти извилистые сосуды, концентрирующиеся возле основания черепа, вдоль спинного мозга, под ребрами грудной клетки и в других местах, были названы чудесной сетью за необычный вид, Чудесную сеть подразделяют на артериальную, венозную и смешанную.
Функциональная роль чудесной сети окончательно не выяснена. Она может служить чем-то вроде резервуара для хранения крови, который быстро наполняется или опорожняется, регулируя, таким образом, кровяное давление при быстром всплытии или погружении. Велика роль этого образования в кровоснабжении мозга, которое осуществляется не прямо из сонных артерий, а исключительно через околопозвоночную чудесную сеть.
В целом можно сказать, что разные морфологические перестройки, например, венозного русла, направлены на обеспечение одного и того же функционального приспособления — депонирования крови и перераспределения кровотока при нырянии.
Высокогорные обитатели
Наряду с целым рядом особенностей высокогорной среды (резкими перепадами температуры, интенсивной инсоляцией и др.) наиболее существенное влияние на дыхательную функцию организма оказывает пониженное парциальное давление кислорода, что вызывает гипоксические реакции. На высоте, превышающей 2-3 тыс. м над уровнем моря Ро2, оказывается в той части кривой диссоциации оксигемоглобина, где уменьшается насыщение крови кислородом. На вершине Эвереста (8848 м над уровнем моря) Ро2 во вдыхаемом воздухе составляет всего лишь 43 мм рт. ст. У людей, впервые попавших на большую высоту, часто наблюдаются головные боли, слабость, головокружение, сильное сердцебиение, тошнота, потеря аппетита и бессонница.. Это связано с развитием острой горной болезни. Чтобы ее избежать, человеку требуется акклиматизация к высокогорью, длящаяся месяцы и годы, а также включение дополнительных физиологических механизмов, компенсирующих недостаточный транспорт кислорода тканям.
Среди позвоночных, адаптированных к условиям высокогорья, есть представители различных классов: амфибии, рептилии, птицы, млекопитающие. Так, некоторых млекопитающих можно встретить в горах на высоте до 6-7 км, т. е. при значениях Ро2, уменьшенных в два-три раза по сравнению с нормальными. Полет птиц наблюдали на высоте 8-10 тыс. м. Более 15 млн человек живет в горах выше 3050 м над уровнем моря, а в Андах поселения встречаются даже на высотах более 4900 м.
Какие же физиологические механизмы позволяют обитателям гор утилизировать малое количество кислорода и вести нормальный образ жизни? По-видимому, адаптация к низкому Ро2 осуществляется в результате повышенной способности извлекать кислород из атмосферы, что определяется целым рядом особенностей функционирования общей газотранспортной системы организма и, в первую очередь, такого ее отдела, как кровеносная система. Прежде всего, это связано с объемом и составом крови. У равнинного жителя объем крови составляет 4-6 л, на Памире он больше на 31,5%, а в Андах на 26%. Общая масса циркулирующих в крови эритроцитов у горцев на 64-83% больше, чем у людей, живущих на равнине. Аналогично меняется и общее содержание гемоглобина в циркулирующей крови. У горцев Памира оно увеличено на 60%, а у жителей Анд на 64% по сравнению с жителями равнин, Таким образом, способность крови транспортировать кислород возрастает, что и требуется в условиях высокогорья. Кроме того, срок жизни эритроцитов у высокогорных обитателей резко отличается от такового у равнинных. У горцев срок жизни эритроцитов почти вдвое меньше, значит, их круговорот и новообразование резко повышены. Следовательно, костный мозг функционирует очень интенсивно, что компенсирует быстрое разрушение эритроцитов. Отмечено также повышенное сродство гемоглобина к кислороду у обитателей горных местностей. Важной особенностью высокогорных жителей является увеличенное содержание миоглобина и его перераспределение в разных группах мышц. Миоглобина очень много в диафрагме, мышцах ног и в сердечной мышце, т. е. именно в тех отделах, которые особенно важны для жизни в горах. Таким образом, для достижения высокой степени адаптации организмов к недостатку кислорода необходимы увеличение общего объема крови, особенности структуры и малая продолжительность жизни эритроцитов, повышенное сродство гемоглобина к кислороду и способность тканей утилизировать кислород с помощью миоглобина.
У горных обитателей адаптивные изменения затронули кроме кровеносной и другие отделы газотранспортной системы организма. Важной компенсаторной реакцией на большой высоте служит гипервентиляция легких. При подъеме в горы первая реакция кардиореспираторной системы на гипоксию проявляется, когда Ро2 вдыхаемого воздуха снижается, до 125-135 мм рт. ст., (что соответствует высоте 1,5-2 тыс. м над уровнем моря). Вследствие возбуждения артериальных хеморецепторов возрастает легочная вентиляция, учащаются сокращения сердца. Эти сдвиги достигают максимума при Ро2 60-70 мм рт. ст. (высота около 7 тыс. м). Следует отметить, что компенсаторного повышения вентиляции легких недостаточно для поддержания нормального напряжения кислорода в альвеолярном газе и артериальной крови, поэтому такой сдвиг можно рассматривать как дополнительное приспособление к горным условиям. Подтверждением этому может служить тот факт, что у высокогорных животных (грызунов, копытных и др.) не обнаружено никаких, специфических особенностей регуляции дыхания. Реакции легочной вентиляции на гипоксию у горных коз, овец, яков и быков оказались такими же, как у родственных им обитателей равнин. Аналогичные результаты были получены для горных жителей (Анды, Гималаи, высота 3-4 тыс. м и более). У них практически не наблюдалось реакции дыхания н





