Реакции субстратного фосфорилирования.
1)реакция 1,3-дифосфоглицерата, содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной кислоты-образуется молекула АТФ 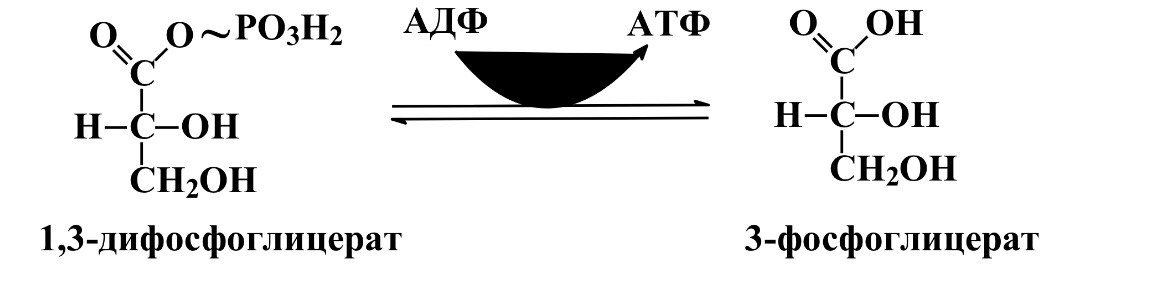
2)реакция субстратного фосфорилирования АДФ с образованием енольной формы пирувата и АТФ, протекающая под действием пируваткиназы 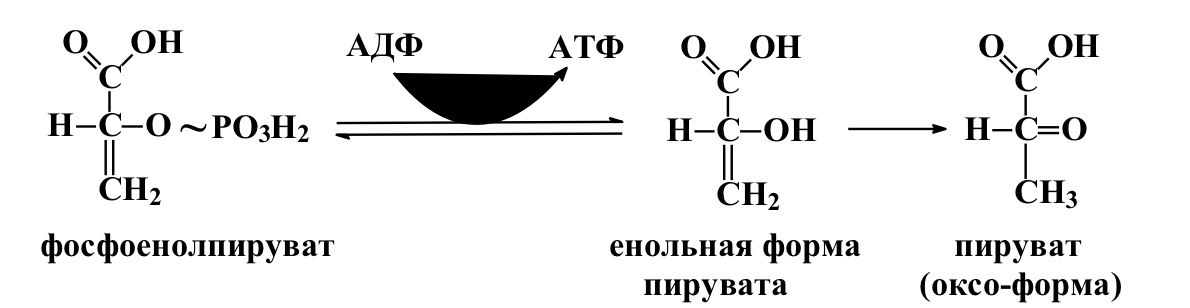
3. Пути и неферментативные реакции образования активных форм кислорода. «Утечка» электронов из неферментативных реакций.
1) Утечка электронов из ЦПЭ на кислород является основным путем образования активных форм кислорода в большинстве клеток:
Пример: В цепи окислительного фосфорилирования Q принимая 1 электрон превращается в свободный радикал семихинон НQ ∙, который при реоксигенации ишемических тканей может непосредственно взаимодействовать с кислородом, образуя супероксидный анион-радикал: HQ· + O2 → Q+ О ∙ 2 + H+;


3) Утечка с цепи микросомального окисления

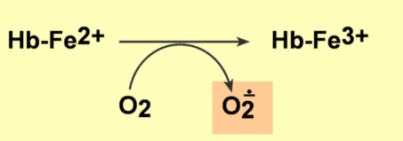
4)Окисление Fe(2+) до Fe(3+) в гемме происходит случайно или под действием токсинов (нитраты, нитриты, нафталин, сульфаниламиды). Окисленная форма гемоглобина (метгемоглобин) не способна переносить O2
Билет 3
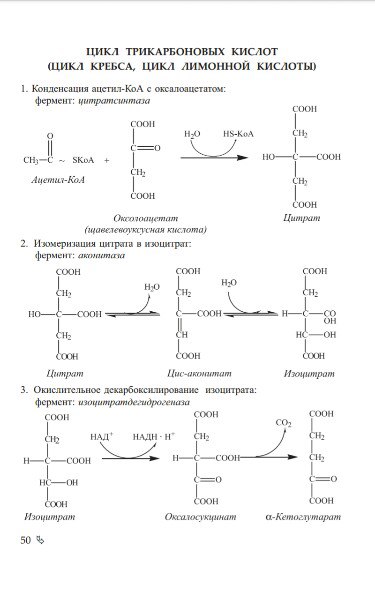
НАПИШИТЕ ПОСЛЕДОВАТЕЛЬНОСТЬ РЕАКЦИЙ ЦИКЛА ТРИКАРБОНОВЫХ
КИСЛОТ. ОТМЕТЬТЕ ЗНАЧЕНИЕ РЕАКЦИЙ ДЕГИДРИРОВАНИЯ И РЕАКЦИИ
СУБСТРАТНОГО ФОСФОРИЛИРОВАНИЯ.

Реакции дегидрирования происходящие в ЦТК служат источником атомов водорода (НАДН2) для дыхательной цепи.
За счет субстратного фосфорилирования образуется энергия в виде ГТФ
ДАЙТЕ ХАРАКТЕРИСТИКУ ПРОЦЕССУ ОКИСЛИТЕЛЬНОГО
МЕХАНИЗМ ПРОЦЕССА, СТАДИИ, КОНЕЧНЫЕ ПРОДУКТЫ. НАПИШИТЕ
СООТВЕТСТВУЮЩИЕ РЕАКЦИИ
Перекисное окисление липидов и представляет собой цепную реакцию, в которой образование одного свободного радикала стимулирует образование других свободных радикалов. В
результате из полиеновых жирных кислот (R) образуются их гидроперекиси (ROOH).
Стадии перекисного окисления липидов
1) Инициация: образование свободного радикала (L•)
Инициирует реакцию чаще всего гидроксильный радикал, отнимающий водород от СН2-групп
полиеновой кислоты, что приводит к образованию липидного радикала.
2) Развитие цепи:
L • + О2 → LOO •
LOO• + LH → LOOM + LR•
Развитие цепи происходит при присоединении О2, в результате чего образуется
липопе-роксирадикал LOO• или пероксид липида LOOH.
ПОЛ представляет собой свободнорадикальные цепные реакции, т.е. каждый образовавшийся радикал инициирует образование нескольких других.
3) Разрушение структуры липидов
4) Обрыв цепи - взаимодействие радикалов между собой:
LOO• + L• → LOOH + LH
L• + vit E → LH + vit E•
vit E• + L• → LH + vit Еокисл.
Конечные продукты перекисного окисления полиеновых кислот - малоновый диальдегид и
гидропероксид кислоты.
Билет №4
1. Опишите строение пируватдегидрогеназного комплекса. Как регулируется пируватдегидрогеназный комплекс? Дайте характеристику витаминов и коферментов, используемых в процессе.
Функция
1. Принимает электроны от НАДН и передает их на коэнзим Q (убихинон).
2. Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
II комплекс
Включает в себя ФАД-зависимые ферменты (напр. сукцианатдегидрогеназа ), расположенные на внутренней мембране.
Функция
1. Восстановление ФАД в окислительно-восстановительных реакциях.
2. Обеспечение передачи электронов от ФАДН2 на железосерные белки внутренней мембраны митохондрий. Далее эти электроны попадают на коэнзим Q.
III комплекс
Функция
1. Принимает электроны от коэнзима Q и передает их на цитохром с.
2. Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
IV комплекс,
цитохром с – кислород-оксидоредуктаза
IV комплекс – цитохромоксидаза или комплекс цитохромов аа3.
Функция
1. Принимает электроны от цитохрома с и передает их на кислород с образованием воды.
2. Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
V комплекс
фермент АТФ-синтаза
Состоит из множества белковых цепей, подразделенных на две большие группы:
субъединица Fо – ее функция каналообразующая, по ней выкачанные наружу протоны водорода устремляются в матрикс.
субъединица F1 – ее функция каталитическая. Она, используя энергию протонов, синтезирует АТФ.
3. Напишите ферментативные реакции образования супероксид анион-радикала, активных форм азота и хлора. Какова дальнейшая судьба и роль этих активных форм?
а) образование супероксид анион-радикала

не обладает сильными окислительными свойствами, но представляет большую опасность, поскольку является источником образования более активных АФК
б) образование активных форм хлора

в) образование активных форм азота

Билет 5
1. Охарактеризуйте цикл трикарбоновых кислот. Укажите его функции и энергетическое значение. Дайте характеристику витаминов и коферментов, используемых в процессе.
ЦТК – заключительный этап катаболизма, в котором углерод ацетильного остатка ацетил Ко-А окисляется до 2 молекул СО2. Атомы водорода, освобождающиеся в ОВР, доставляются в ЦПЭ при участии NAD- и FAD-зависимых дегидрогеназ, в результате чего происходят синтез воды и окислительные фосфорилирование АДФ. Связь между атомами углерода в ацетил Ко-А устойчива к окислению. В условиях организма окисление ацетильного остатка происходит в несколько этапов, образующих циклический процесс из 8 реакций.

Каждый оборот цикла лимонной кислоты сопровождается синтезом 11 молекул АТФ путем окислительного фосфорилирования. Одна молекула АТФ образуется путем субстратного фосфорилирования. В итоге на каждый ацетильный остаток, включенный в ЦТК, образуется 12 молекул АТФ.
Значение этого процесса в том, что он является главным генератором водорода для цепей транспорта электронов, то есть именно он в основном обеспечивает их функционирование, а, следовательно, и синтез АТФ. Локализован цикл Кребса в матриксе митохондрий. Этот процесс является интегративным – он объединяет пути катаболизма углеводов, белков и липидов: в ходе распада всех этих соединений появляется ацетил Ко-А, который и расщепляется до конца в ЦТК.
Ферменты и коферменты:
1 реакция
Пируватдегидрогеназа (Е1), кофермент – тиаминдифосфат, функция – окислительное декарбоксилирование пировиноградной кислоты,
2 и 3 реакции
Дигидролипоат-ацетилтрансфераза (Е2), кофермент – липоевая кислота, функция – перенос ацетильной группы на HS-КоА и образование ацетил-SКоА
4 и 5 реакции
Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, функция – регенерация окисленной формы липоамида, образование НАДН+Н+ и возвращение комплекса в исходное состояние.
Регуляторными являются следующие ферменты ЦТК: цитратсинтаза (ингибируется НАДН и АТФ), изоцитратдегидрогеназа (ингибируется НАДН и АТФ, активируется АДФ), сукцинатдегидрогеназа (ингибируется ЩУК, активируется сукцинатом, фумаратом, Н3РО4)
2. Дайте характеристику процессу окислительного фосфорилирования. Поясните механизм сопряжения окисления и фосфорилирования.
Окислительным фосфорилированием называют синтез АТФ путем фосфорилирования АДФ за счет энергии трансмембранного электрохим потенциала, возникающего при освобождении энергии электронами окисленного субстрата в процессе миграции этих электронов по дыхательной цепи к вдыхаемому кислороду. То есть окисл фосфорилирование – это синтез АТФ, связанный с переносом электронов по дыхательной цепи, это синтез АТФ за счет энергии окисления субстрата. Происходит в аэробных условиях
Основные этапы окислительного фосфорилирования:
1. Образующиеся в реакциях катаболизма НАДН и ФАДН2 передают атомы водорода (т.е. протоны водорода и электроны) на ферменты дыхательной цепи.
2. Электроны движутся по ферментам дыхательной цепи и теряют энергию.
3. Энергия электронов используется на выкачивание протонов Н+ из матрикса в межмембранное пространство.
4. В конце дыхательной цепи электроны попадают на кислород и восстанавливают его до воды.
5. Протоны Н+ стремятся обратно в матрикс и проходят через АТФ-синтазу.
6. При этом они теряют энергию, которая используется для синтеза АТФ.
Под сопряжением окисления с фосфорилированием понимают превращение энергии электронов окисленного субстрата, проходящих по дыхательной цепи, в промежуточную форму – в энергию трансмембранного потенциала с последующим использованием ее для фосфорилирования АДФ, то есть для синтеза АТФ. Протонный градиент создается путем выталкивания ионов водорода в межмебранное пространство в 3 участках дых цепи: при переходе электронов с ФМНН2 через FeS-белок на KoQ, при переходе электронов с KoQH2 через FeS-белок на цитохром Ci при переходе электронов от цитохрома, а через цитохром а3 к кислороду. Эти участки цепи транспорта электронов обозначают как пункты сопряжения дыхания с фосфорилированием.
3. Напишите ферментативные реакции образования пероксида водорода. Какова дальнейшая судьба и роль пероксида водорода?
Пероксид водорода образуется с помощью фермента супероксидазы из:
а) диоксида 2∙О2- + 2H+ → Н2О2 + О2
б) протонированной формы диоксида 2 НО2∙ → Н2О2 + О2
Пероксид водорода не является радикалом и, будучи нейтральной молекулой, легко проникает через гидрофобные мембраны. Токсические эффекты перекиси водорода проявляются при концентрации более 5х10^-5, которая наблюдается в очагах воспаления благодаря активации фагоцитирующих клеток. Пероксид водорода занимает особое место в генерации АФК в лейкоцитах для уничтожения инфицирующих микроорганизмов.
Н2О2 в присутствии миэлопероксидазы и перксидазы эозинофилов служит источником гипогалогенидов (например, HClO), которые тоже токсичны и подавляют микрофлору:
HCl + Н2О2 → HClO + H2O
В организме пероксид водорода постоянно разрушается за счет ферментов каталазы
(2Н2О2 → 2Н2О +О2) и пероксидазы (SH2 + Н2О2 → S + 2H2O).
Свободно проникая внутрь клетки, перокисид водорода взаимодействует с ионами металлов (железо, медь), в результате чего образуется гидроксид-радикал (гидроксильный радикал) НО∙:
Н2О2+ Cu+ → Cu2+ + ОН- + НО∙
Билет 6
1.Напишите последовательность реакций цикла трикарбоновых кислот. Поясните анаболические функции цикла трикарбоновых кислот. Отметьте анаплеротические реакции и их значение. 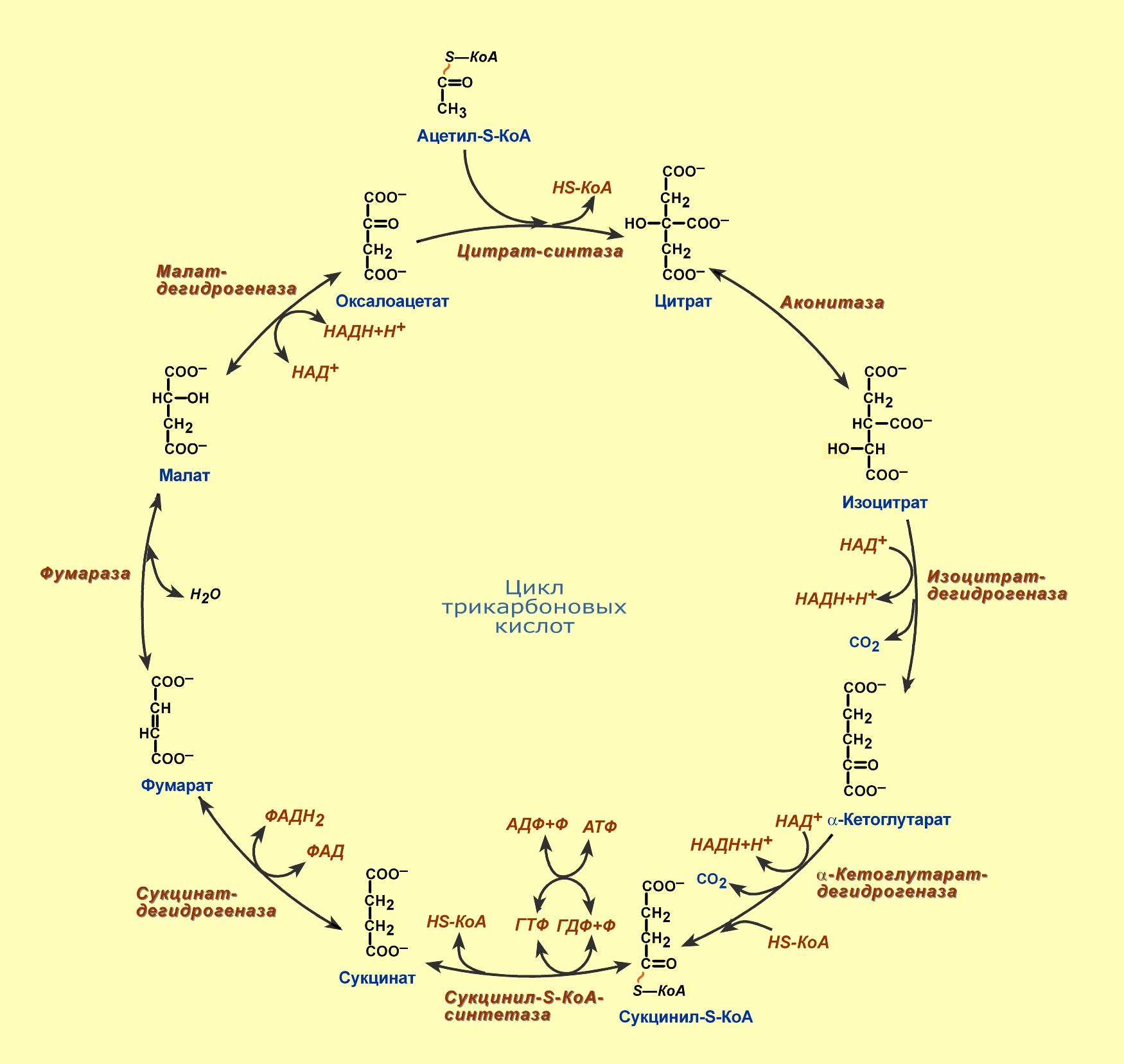

Анаплеротические пути
Промежуточные соединения цикла трикарбоновых кислот, покинувшие цикл и задействованные в синтезе различных соединений, замещаются за счёт специальных анаплеротических реакций. В нормальных условиях реакции, в ходе которых промежуточные соединения цикла вовлекаются в другие метаболические пути, и реакции, возмещающие их уход, находятся в динамическом равновесии, поэтому концентрация промежуточных соединений цикла трикарбоновых кислот поддерживается постоянно.
Ниже в таблице представлены важнейшие анаплеротические реакции:
| Реакция
| Фермент
| Ткань/организм
| |
| пируват + НСО3− + ATP ⇌ оксалоацетат + ADP + Pi
| пируваткарбоксилаза
| печень, почки
|
|
| фосфоенолпируват + СО2 + GDP ⇌ оксалоацетат + GTP
| фосфоенолпируваткарбоксикиназа
| сердце,скелетные мышцы
|
|
| фосфоенолпируват + НСО3− ⇌ оксалоацетат + Pi
| фосфоенолпируваткарбоксилаза
| высшие растения,дрожжи, бактерии
|
|
| пируват + НСО3− + NAD(P)H ⇌ малат + NAD(P)+
| малик-энзим
| широко распространена среди эукариот и бактерий
|
|
Регуляция ОФ
Работа дыхательных ферментов регулируется с помощью эффекта,который получил название дыхательный контроль. Дыхательный контроль – это прямое влияние электрохимического градиента на скорость движения электронов по дыхательной цепи. Величина градиента напрямую зависит от соотношения АТФ и АДФ, количественная сумма которых в клетке постоянна
[АТФ] + [АДФ] = const
Возрастание протонного градиента возникает при отсутствии АДФ и накоплении АТФ, т.е. когда АТФ-синтаза лишена своего субстрата и ионы Н+ не проникают в матрикс митохондрии.
При этом ингибирующее влияние градиента усиливается и продвижение электронов по цепи замедляется.
Снижение протонного градиента возникает при наличии АДФ и прохождении ионов Н+ через канал Fо в матрикс. В этом случае:
• протонный градиент снижается,
• скорость потока электронов возрастает,
• усиливается выкачивание ионов Н+ в межмембранное пространство,
• увеличивается окисление НАДН и ФАДН2 (как источников электронов),
• снимается ингибирующее влияние НАДН на цикл лимонной кислоты и ПВК-дегидрогеназу.
Билет 10
3) Назовите и дайте характеристику неферментативным антиоксидантам небелковой природы. Напишите соответствующие реакции.
Неферментативные компоненты антиоксидантной системы:
1. Витамины Е (токоферол) и А (ретинол), витамин С, каротиноиды и изопренои-
ды, которые находятся в составе клеточных мембран.
2. Церулоплазмин - белок плазмы крови, который принимает участие в транспорте меди
3. Мочевая кислота.
Механизм действия этих компонентов:
они принимают неспаренные электроны от активных форм кислорода, при этом образуется радикал антиоксиданта, который малоактивен. Таким образом неферментативные компоненты антиоксидантной системы — это перехватчики неспаренных электронов. 
(!) К неферменентативным антиоксидантам НЕбелковой природы относят низмолекулярные соединения, которые относятся ко второй линии защиты. Т.е. витамин Е и С, каротиноиды, изопреноиды и так называемых непищевые антиоксиданты, например, фенольные и полифенольные соединения.
• Витамин Е, встраиваясь в фосфолипидный бислой мембран, выполняет антиоксидантную функцию, т.е. препятствует развитию свободнорадикальных реакций. При этом: Защищает витамин А от окисления, что способствует проявлению ростстимулирующей активности витамина А.
Защищает жирнокислотные остатки мембранных фосфолипидов, и, следовательно, любые клеточные мембраны от перекисного окисления
• Витамин А. Благодаря наличию двойных связей в изопреновой цепи витамин осуществляет нейтрализацию свободных кислородных радикалов, особенно эта функция проявляется у каротиноидов.
• Витамин С. Антиоксидантная роль:
• сбережение витамина Е,
• лимитирование свободнорадикальных реакций в делящихся тканях,
• ограничивает воспаление.
2) Каким образом регулируется окислительное фосфорилирование? Назовите и охарактеризуйте ингибиторы окислительного фосфорилирования.
Работа дыхательных ферментов регулируется с помощью эффекта, который получил название дыхательный контроль.
Дыхательный контроль – это прямое влияние электрохимического градиента на скорость движения электронов по дыхательной цепи (величину дыхания). В свою очередь, величина градиента напрямую зависит от соотношения АТФ и АДФ, количественная сумма которых в клетке постоянна ([АТФ] + [АДФ] = const).
Возрастание протонного градиента возникает при исчерпании запасов АДФ и накоплении АТФ, т.е. когда АТФ-синтаза лишена своего субстрата и ионы Н+ не проникают в матрикс митохондрии. При этом ингибирующее влияние градиента усиливается и продвижение электронов по цепи замедляется. Ферментные комплексы остаются в восстановленном со- стоянии. Следствием является уменьшение окисления НАДН и ФАДН2 на I и II комплексах и замедление катаболизма в клетке.
Снижение протонного градиента возникает при активной работе АТФ-синтазы (т.е. прохождении ионов Н+ в матрикс). В этом случае градиент снижается, поток электронов воз- растает, в результате повышается выкачивание ионов Н+ в межмембранное пространство. Ферментные комплексы I и II усиливают окисление НАДН и ФАДН2 (как источников элек- тронов) и снимается ингибирующее влияние НАДН на цикл лимонной кислоты и пируватдегидрогеназный комплекс – активируются реакции катаболизма углеводов и жиров, как источника энергии.
• Ингибиторы блокируют ферменты дыхательной цепи и замедляют движение электронов от НАДН и ФАДН2 на кислород.
Выделяют три основных группы ингибиторов:
действующие на I комплекс, например, амитал, ротенон, прогестерон,
действующие на III комплекс, например, антимицин А,
действующие на IV комплекс, например, сероводород (H2S), угарный газ (СО), циа-
ниды (-CN).
1) Охарактеризуйте роль витаминов и их коферментых форм в общих путях катаболизма. Напишите примеры реакций, в которых они принимают участие.
Общий путь катаболизма включает 2 процесса: окислительное декарбоксилирование пирувата и цикл трикарбоновых кислот Кребса. 
- Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиамин- дифосфат, катализирует 1-ю реакцию. (Витамин В1, тиамин)
- Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции.
- Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими фер- ментами, в работе комплекса принимают участие коэнзим А и НАД. (Витамин В2, рибофлавин)
Цикл Кребса: роль витаминов
В цикле лимонной кислоты (Цикл Кребса: метаболическая карта) выполняют специфические функции четыре водорастворимых витамина группы B.
1) Рибофлавин входит в состав флавинадениндинуклеотид а (FAD), который является кофактором альфакетоглутаратдегидрогеназного комплекса и сукцинатдегидрогеназы.
2) Ниацин входит в состав никотинамидадениндииуклеотид а (NAD), который является коферментом трех дегидрогеназ цикла: изоцитратдегидрогеназы, альфа-кетоглутаратдегидрогеназы и малатдегидрогеназы.
3) Тиамин (витамин B1) входит в состав тиаминдифосфата, который является коферментом альфа-кетоглутаратдегидрогеназы.
4) Пантотеновая кислота входит в состав кофермента A, который является кофактором, связывающим "активные" ацильные остатки, например, в ацетил-CoA или сукцинил-CoA.
11 Билет
1. Напишите суммарную и отдельные реакции окислительного декарбоксилирования пирувата.
Пировиноградная кислота (ПВК, пируват) является продуктом окисления глюкозы и некоторых аминокислот. В анаэробных условиях она восстанавливается до молочной кислоты. В аэробных условиях происходит ее окислительное декарбоксилирование до уксусной кислоты, переносчиком которой служит коэнзим А.
Суммарное уравнение реакции отражает декарбоксилирование пирувата, восстановление НАД до НАДН и образование ацетил-S-KoA.

Превращение состоит из пяти последовательных реакций, осуществляется мультиферментным комплексом, прикрепленным к внутренней митохондриальной мембране со стороны матрикса. В составе комплекса насчитывают 3 фермента и 5 коферментов:
Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиамин-дифосфат, катализирует 1-ю реакцию.
Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кисло-та, катализирует 2-ю и 3-ю реакции.
Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реак-ции.
Помимо указанных коферментов, которые прочно связаны с соответствующими фер-ментами, в работе комплекса принимают участие коэнзим А и НАД.
Суть первых трех реакций сводится к декарбоксилированию пирувата (катализируется пируватдегидрогеназой, Е1), окислению пирувата до ацетила и переносу ацетила на коэн-зим А (катализируется дигидролипоамид-ацетилтрансферазой, Е2).

Оставшиеся 2 реакции необходимы для возвращения липоевой кислоты и ФАД в окисленное состояние (катализируются дигидролипоат-дегидрогеназой, Е3). При этом образуется НАДН.

2. Охарктеризуйте пути синтеза АТФ и виды фосфорилирования.Напишите реакции субстратного фосфорилирования.
В организме АТФ синтезируется путём фосфорилирования АДФ:
АДФ + H3PO4 + энергия → АТФ + H2O.
Фосфорилирование АДФ возможно двумя способами:
А)субстратное фосфорилирование
Б)окислительное фосфорилирование
Субстратное фосфорилирование. Этот способ связан с передачей макроэргического фосфата или энергии макроэргической связи какого-либо вещества (субстрата) на АДФ. К таким веществам относятся метаболиты гликолиза (1,3-дифосфоглицериновая кислота, фосфоенолпируват), цикла трикарбоновых кислот (сукцинил-S-КоА) и креатинфосфат.
Окислительное фосфорилирование (3 этап) – окисляются НАДН и ФАДН2, полученные в реакциях катаболизма глюкозы и жирных кислот. При этом ферменты внутренней мембраны митохондрий обеспечивают образование основного количества клеточного АТФ из АДФ


3. Назовите и дайте характеристику ферментам глутатионовой системы антиоксидантной защиты. Напишите соответствующие реакции
Глутатионпероксидаза также восстанавливает гидропероксиды липидов.
Глутатионпероксидаза. SH-группа цистеина, входящего в состав глутатиона, может отдавать всего 1 атом водорода, а для пероксидазной реакции необходимы 2 атома. Поэтому молекулы глутатиона работают парами.
Глутатионпероксидаза присутствует во всех клетках организма и разлагает пероксид водорода. Содержит селеноцистеин в активном центре.
2 Глутатион-SH + Н2О2 → Глутатион-SS + 2Н2О
Реакция, катализируемая глутатионпероксидазой:
2Н2О2 + 2Г-SH → H2O + Г-S-S-Г
Регенерация глутатиона идет с участием НАДФН, катализирует ее фермент глутатионредуктаза.
Г-S-S-Г + НАДФН(Н+) → 2Г-SH + НАДФ
Глутатион постоянно поддерживается в восстановленном состоянии в эритроцитах, где он служит для защиты гема гемоглобина от окисления.

Билет 12
1.Цикл трикарбоновых кислот – система реакций, приводящая к полному окислению двухуглеродного ацильного остатка. Является общим конечным путем окисления белков, жиров и углеводов.
Локализован в митохондриях.
В ходе одного полного цикла происходит:
1. полное окисление ацильного остатка до двух СО2.
2. Образование 3 молекул восстановленного NADH+H и одной мол. FADH2
3. Синтез 1 мол. ГТФ.
За 1 цикл синтез. 12 мол АТФ. 9 АТФ от 3 NADH+H. 2 АТФ от FADH2 1ГТФ=1АТФ
Функции: Полное окисление двухуглеродного ацильного остатка. Выделившуюся энергию преобразовать в АТФ.
Коферменты:
Кофермент А (коэнзим А, КоА) — кофермент ацетилирования. При расщеплении тиоэфирной связи освобождается коэнзим А и выделяется значительная тепловая энергия. Рав- новесие в реакции сильно сдвинуто вправо.
5 коферментов: тиаминдифосфат (ТДФ), липоамид, ФАД, НАД+ и СоА.
Витамины: Рибофлавин (В2) входит в состав флавинадениндинуклеотида (ФАД), который является коферментом α–кетоглутаратдегидрогеназного комплекса и сукцинатдегидрогеназы;
Никотинамид (РР) включен в структуру никотинамидаденин-динуклеотида (НАД+), служащего коферментом трех энзимов: изоцитрат-, малатдегидрогеназы и α–кетоглутаратдегидрогеназном комплекс;,
Тиамин (В1) в виде тиаминдифосфата выполняет функцию кофермента в α–кетоглутаратдегидрогеназном комплексе;
Пантотеновая кислота (В3) – составная часть коэнзима А, связывающего активные ацильные остатки, например, в ацетил-КоА или сукцинил-КоА;
Липоамид – витаминоподобное соединение служит коферментом α–кетоглутаратдегидрогеназы
2. Дыхательная цепь состоит из переносчиков электронов, сгруппированных в 4 ферментативных комплекса, и ферментов. В Геме Fe 
Функции: 1 комплекс восстановление убихинона.
2 комплекс убихинон востановляется в убихинол.
3 комплекс окисление убихинола перенос электронов на цитохром с.
4 комплекс перенос электрона на кислород, образование Воды Гемы: железо(а,а3) и медь(а3).
5 комплекс синтез АТФ

3.местное образование свободных радикалов иммунными клетками (фермент НАДФН-оксидаза нейтрофилов) может быть важным фактором в борьбе организма с
бактериями и защите его от инфекции.
Свободный радикал — частица, обладающая одним или несколькими
неспаренными электронами Свободные радикалы и различные активные
формы кислорода могут образовываться либо при нормальных, основных ме-
таболических процессах, либо под воздействием внешних источников.
Гидроксид-радикал является главным инициатором пероксидного
окисления липидов (ПОЛ). Т.Е. метаболизм жиров(хз).
Пероксид водорода занимает особое место в генерации АФК в лейко-
цитах для уничтожения инфицирующих микроорганизмов. Н2О2 в присут-
ствии миэлопероксидазы и перксидазы эозинофилов служит источником ги-
погалогенидов (например, HClO), которые тоже токсичны и подавляют мик-
рофлору.
Билет 13
Дисфункции митохондрий
Известно, что митохондрии играют доминирующую роль в обеспечении тканей организма энергией, генерации супероксид - радикала, реализации механизмов программированной гибели клетки и участвуют в депонировании внутриклеточных
ионов Са2+. Это определяет центральную роль митохондрий в развитии многих патологических процессов.
Механизмы:
Приобретенные нарушения митохондриальных функций при различных патологических процессах, возникают преимущественно как следствие повреждения митохондриальных структур под влиянием свободно-радикальных процессов, активизации механизмов деградации митохондрий, стимуляции апоптогенных сигналов, увеличения концентрации ионов кальция в цитоплазме клеток органов-мишеней.
Наиболее распространенными факторами являются нарушения липидного обмена, гипергликемия, формирование белковых агрегатов в клетках, ишемии\реперфузии, токсических повреждениях. Существуют наследственные заболевания, ассоциированные с врожденными мутациями в митохондриальном геноме.
Основным следствием нарушения энергетического обмена в митохондриях является нарушение синтеза АТФ и подавление β-окисления липидов. Показано, что существует определенный пороговый уровень снижения внутриклеточного содержания АТФ, после достижения которого, начинается резкое подавление любых энергозатратных процессов в клетке.
Известно, что нарушение переноса электронов между компонентами дыхательной цепи сопровождается генерацией митохондриями супероксидного радикала. Он способствует активизации процессов перекисного окисления липидов, оксидативных повреждений белков и нуклеиновых кислот. Увеличение митохондриями активных форм кислорода может оказать на клетку не только повреждающий, но и иррегуляторный эффект.
Супероксиддисмутаза
Супероксидный анион спонтанно или при участии фермента супероксиддисмутазы превращается в пероксид водорода:
О2- + О2- + 2Н+ → Н2О2 + О2.
Миелопероксидаза
Под действием миелопероксидазы, проникающей в фагосому при её слиянии с лизосомой, из пероксидов в присутствии галогенов (йоди-дов и хлоридов) образуются дополнительные токсичные окислители - гипойодид и гипохлорид.
Н2О2 + Cl- + H+ → НОС1 + H2O.
Все эти молекулы являются сильными окислителями и оказывают бактерицидное действие. Резкое увеличение потребления кислорода фагоцитирующей клеткой называется "респираторным взрывом"
1.Повышение потребление глюкозы и ее окисления по пентозофосфатному пути с образованием НАДФН.
2. Повышение потребления кислорода в 20-40 раз.
3. Активация НАДФН-оксидазы и генерация супероксид анион-радикала.
4. Активация миелопероксидазы и генерация гипохлорита.
Активные формы кислорода инициируют свободнорадикальные реакции, разрушающие липиды клеточных мембран поглощённых фагоцитами бактерий.
Наследственная недостаточность NADP-оксидазы, обусловленная дефектом одного из генов этого ферментного комплекса, приводит к хроническому гранулематозу. В результате дефекта фермента фагоциты больных не способны продуцировать супероксидный кислородный радикал и пероксид водорода и поэтому не могут быстро разрушать фагоцитированные клетки бактерий и грибов. Некоторые устойчивые микроорганизмы остаются жизнеспособными внутри фагоцитов, и их антигены вызывают в месте скопления фагоцитов клеточный иммунный ответ и формирование гранулём. Наиболее часто встречается сцепленная с Х-хромосомой форма этого заболевания, связанная с дефектом гена одной из полипептидных цепей комплекса, локализованного на коротком плече Х-хромосомы.
Образование реакционноспособных метаболитов азота. Бактерицидное действие в макрофагах оказывает и оксид азота (NO). Оксид азота в этих клетках образуется, так же как и в других, под действием фермента NO синтазы из аргинина (см. раздел 9). Активность NO синтазы в макрофагах заметно повышается при фагоцитозе в присутствии γ-интерферона и фактора некроза опухолей. Супероксид-анион образует с NO соединения, обладающие большими бактерицидными свойствами, чем сам NO:
NO + О2- → ONOO- → ОН* + NO2.
Билет 28
Вопрос 1. Опишите строение пируватдегидрогеназного комплека. Как регулируется пируватдегидрогеназный комплекс? Дайте характеристику витаминов и коферментов, используемых в процессе.
В составе комплекса насчитывают 3 фермента и 5 коферментов:
Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиамин- дифосфат, катализирует 1-ю реакцию.
Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции.
Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реак- ции.
Помимо указанных коферментов, которые прочно связаны с соответствующими фер- ментами, в работе комплекса принимают участие коэнзим А и НАД.


Регуляция ПВК
Осуществляется различными способами:
доступностью субстратов, ингибированием продуктами реакциии, аллостерически и путем ковалентной модификации.
1) Ковалентная модификация
В пируватдегидрогеназном комплексе имеются еще 2 вспомогательных фермента – ки- наза и фосфатаза, участвующие регуляции активности пируватдегидрогеназы (Е1) путем фосфорилирования и дефосфорилирования.
Киназа активируется при избытке АТФ и продуктов реакции – НАДН и ацетил-SКоА.
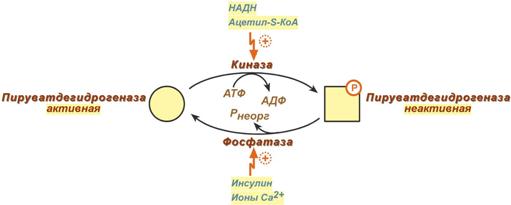
2) Аллостерическая регуляция
ингибиторы - продукты реакции (ацетил-КоА и НАДН) активируют киназу ПДК
активаторы - субстраты реакции (пируват, НАД+, КоА), а также инсулин и ионы Cа2+
Коферменты:
1. Тиаминдифосфат - производное витамина В1 (тиамин)
2. Флавинадениндинуклеотид - производное витамина В2 (рибофлавин)
3. Никотинамидадениндинуклеотид - производное витамина В3 (ниацин)
4. Коэнзим А - производное витамина В5 (фолиевая кислота)
Вопрос 2. Дайте харакеристику митохондриям, их функции и дисфункции. Что такое митохондриальные болезни? Укажите причины развития гипоэнергетических состояний.
Митохондрия ограничена двумя мембранами - гладкой внешней и складчатой внутренней, имеющей очень большую поверхность. Складки внутренней мембраны глубоко входят в матрикс митохондрий, образуя впячивания - кристы. Пространство между внешней и внутренней мембранами обычно называют межмембранным пространством.
Мембраны митохондрий содержат интегральные мембранные белки. Во внешнюю мембрану входят порины, которые образуют поры и делают мембраны проницаемыми для веществ с молекулярной массой до 10 кДа. Внутренняя же мембрана митохондрий ПОЧТИ исключение составляют О2, СО2, Н20. Внутренняя мембрана митохондрий характеризуется необычно высоким содержанием белков (75%). В их число входят транспортные белки-переносчики, ферменты, компоненты дыхательной цепи и АТФ-синтаза.
Функции:
В них синтезируется большая часть необходимого клетке АТФ.
В митохондриях локализованы следующие метаболические процессы: превращение пирувата в ацетил-КоА, катализируемое пируватдегидрогеназным комплексом: цитратный цикл; дыхательная цепь, сопряженная с синтезом АТФ («окислительное фосфорилирование»);
Патологии митохондрий
Передаются только по материнской линии. При делении митохондрий, ДНК случайным образом распределяется между потомками. Мутации мтхДНК могут накапливаться и приводить к тяжелым заболеваниям.
Атрофия зрительного нерва Лебера
Вызвана мутацией участка мтхДНК, который кодирует одну из субъединиц комплекса I. Болезнь проявляется во взрослом возрасте и приводит к потере зрения.
Митохондриаьная энцефаломиопатия, лакоацидоз и инсультоподобные эпизоды (синдром MELAS)
Вызвана мутацией в гене, кодирующем тРНК митохондрий. Эта мутация влияет на транскрипцию мтДНК, в рез-те чего нарушается структура всех комплексов дых. цепи, кроме II, который полностью кодируется ядерным геномом.
Болезнь Ли
Дегенеративное заболевание ЦНС, обычно развивается в раннем возрасте. Чаще всего причиной болезни является мутации участков ядерных геномов, кодирущих компоненты дых. цепи. Однако в некоторых случаях бол





