

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Топ:
Процедура выполнения команд. Рабочий цикл процессора: Функционирование процессора в основном состоит из повторяющихся рабочих циклов, каждый из которых соответствует...
Эволюция кровеносной системы позвоночных животных: Биологическая эволюция – необратимый процесс исторического развития живой природы...
Установка замедленного коксования: Чем выше температура и ниже давление, тем место разрыва углеродной цепи всё больше смещается к её концу и значительно возрастает...
Интересное:
Аура как энергетическое поле: многослойную ауру человека можно представить себе подобным...
Подходы к решению темы фильма: Существует три основных типа исторического фильма, имеющих между собой много общего...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Исследования бинокулярной конкуренции (Binocular rivalry)
Зрительные галлюцинации
ЭЭГ и МЭГ исследования зрительного осознания
Транскраниальная магнитная стимуляция (ТМС)
Выводы
Вопросы для обсуждения
Введение: зрительная информация и зрительное осознание
В исследованиях НКС, посвященных определенному содержанию сознания, зрительное осознание, безусловно, изучают чаще всего. Поэтому здесь мы рассмотрим некоторые из самых показательных экспериментов в сфере нейрональных коррелятов зрительного осознания.
Зрительная информация поступает от глаз в мозг через ядра таламуса (латеральные коленчатые тела) и заканчивает свой путь в первичной зрительной коре (V1), которую еще называют стриатной корой. Оттуда она поступает в сложную сеть областей коры мозга, которые специализируются на обработке различных аспектов зрительной информации. Два основных пути обработки называются вентральным зрительным трактом (он идет от области V1 к экстрастриатной затылочной коре, а затем к височной доле) и дорсальным зрительным трактом (он идет от области V1 к задней париетальной коре (см. рис. 5.3). Вентральный зрительный тракт содержит области, обрабатывающие такие зрительные характеристики, как цвет, форма и тип объекта (например, лицо, животное или место). Таким образом, вентральный тракт, как предполагается, собирает воедино представления о связанных зрительных характеристиках, позволяющие их распознавать и объединять в целостный объект. Дорсальный зрительный тракт связан с пространственным расположением и движением объектов. Считается, что он помогает отслеживать движение объектов в пространстве и управляет сенсомоторным взаимодействием с объектами.
|
|
Возникает вопрос: в каких областях зрительной зоны коры мозга расположены нейрональные корреляты осознаваемого зрительного восприятия? Что происходит в мозге, когда мы видим объект или его элемент или когда зрительная информация входит в наше сознание в последовательной феноменальной форме, в виде красивых, ярких зрительных квалиа, например хроматических цветов? Чтобы исследовать этот вопрос, нужно разработать эксперименты, где в одних условиях зрительная информация не входит в сознание – субъективно мы не видим стимулов, а в других условиях та же самая зрительная информация входит в сознание, и у нас возникает субъективное зрительное восприятие стимула.
Исследование бинокулярной конкуренции
Для экспериментов, позволяющих отделить нейрональные корреляты субъективного зрительного переживания от активности, непосредственно вызванной зрительными стимулами, один подход оказался особенно плодотворным. Это исследования феномена, который называется бинокулярной конкуренцией. Ее можно вызвать, одновременно показывая два разных, зрительно несовместимых стимула, один для левого глаза, а другой – для правого (рис. 9.1). Обычно мозг объединяет информацию от обоих глаз в одно целостное трехмерное изображение, но в этом случае такая интеграция невозможна, потому что два представления не соответствуют друг другу. Скажем, левому глазу мы показываем изображение кролика, а правому глазу – изображение цветка.
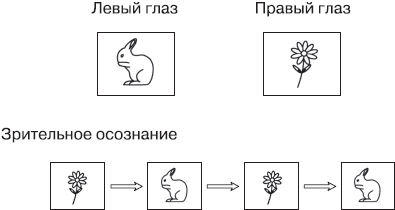
Рис. 9.1. Бинокулярная конкуренция
Левый глаз все время «видит» только кролика, а правый глаз все время «видит» только цветок. Но в зрительном осознании цветок и кролик возникают последовательно, друг за другом, и содержание сознания спонтанно меняется каждые несколько секунд
Сначала мозг пытается объединить оба образа, но когда это не удается, изображения начинают конкурировать друг с другом за доступ в сознание. Таким образом, сначала, в течение нескольких секунд, испытуемый видит цветок и больше ничего, а затем цветок внезапно превращается в кролика. В течение нескольких следующих секунд испытуемый видит кролика, а потом образы снова переключаются. Спонтанная конкуренция между двумя глазами и двумя стимулами может продолжаться довольно долго. Этот феномен дает идеальную возможность исследовать, что происходит в мозге, когда информация оказывается в сознании. Во время эксперимента оба стимула постоянно находятся перед глазами испытуемого – они остаются неизменными. Поэтому испытуемого просят сообщить, когда у него в сознании происходит переключение образов. Например, он может нажимать на кнопку.
|
|
В экспериментах с бинокулярной конкуренцией участвовали и обезьяны, и люди (хотя, как можно предположить, было не так просто научить обезьян сообщать , что они видят!). В экспериментах с участием обезьян активность единственной клетки в зрительной коре мозга регистрировалась с помощью вживленных микроэлектродов: это тонкие иглы, введенные в зрительную кору головного мозга, и острие иглы регистрирует электрическую активность нейронов, расположенных рядом с ней.
Таким образом удавалось точно определить, в каких участках мозга возникает активность, но сигнал отражал только реакцию одного или нескольких нейронов на изменения в субъективном переживании. Итак, исследователи регистрировали реакции клеток, «интересовавшихся» стимулами, которые видела обезьяна, – эти клетки реагировали на наличие или отсутствие физического стимула в поле зрения при отсутствии конкуренции. Некоторые клетки активировались, когда обезьяна видела изображение лица, а другие – когда она видела изображения каких‑то других объектов, при отсутствии других стимулов.
Когда два изображения показывали одновременно, в ситуации конкуренции между ними, результаты показали, что в первичной зрительной коре (V1) лишь для немногих из клеток, активность которых регистрировалась (около 20%), эта активность коррелировала с изменениями в содержании сознания. Когда регистрировалась активность клеток других областей вдоль вентрального зрительного тракта, количество клеток, коррелирующих с изменениями сознания, увеличивалось вдоль всего вентрального тракта. Таким образом, в задней части височной до ли уже 90% клеток, активность которых регистрировалась, «интересовались» стимулами данного типа, при этом их активность коррелировала с изменениями в содержании сознания: они были максимально активны, когда предпочитаемый стимул появлялся в сознании, – в отличие от тех ситуаций, когда он просто находился перед глазами обезьяны и воспринимался мозгом, но не достигал сознания. Активность этих клеток снижалась, когда предпочитаемый стимул исчезал из сознания и в сознании появлялся конкурирующий стимул (Leopold & Logothetis, 1999).
|
|
После столь захватывающих экспериментов на обезьянах подобный эксперимент был проведен с участием людей, но на этот раз активность мозга регистрировалась с помощью фМРТ. Прежде всего, оказалось, что когда испытуемому показывают изображение лица, максимально активируется особая область вентрального тракта, «фузиформная лицевая область», а когда испытуемый видит изображение дома, активируется другая область, «парагиппокампальная область места». Когда и лицо и дом демонстрировали одновременно в условиях конкуренции, уровень активности этих областей коррелировал с содержанием сознания. Когда в сознании появлялся образ лица, активировалась область, отвечающая за восприятие лиц, а когда в сознании возникало изображение дома, активировалась область, отвечающая за восприятие места. Таким образом, возникает впечатление, что области, которые специализируются на обработке определенных типов объектов в мозге человека, влияют на зрительное осознание объектов того же типа (Kanwisher, 2001).
Зрительные галлюцинации
Независимые доказательства одного и того же явления были получены в ходе экспериментов, где наличие или отсутствие определенного содержания в зрительном осознании возникало спонтанно, вообще без каких бы то ни было стимулов. Неврологическое расстройство под названием «синдром Чарльза Боннета» характеризуется спонтанными зрительными галлюцинациями в виде объектов, лиц, людей или других зрительных иллюзий. Эти галлюцинации могут быть настолько яркими и реалистичными, что их легко перепутать с реальностью. Однако каждый больной обычно видит особые, но всегда одинаковые образы, и поэтому легко учится распознавать свои галлюцинации и не путать их с реальностью (Fychte и др., 1998).
|
|
Во время эксперимента проводилась запись активности мозга с помощью фМРТ. В контрольных условиях у пациента не было галлюцинаций, а в экспериментальных условиях содержание галлюцинаций явно присутствовало в зрительном осознании. Результаты показали, что наличие галлюцинации коррелирует с активностью определенных областей вдоль вентрального зрительного тракта.
В целом доказательства, полученные в ходе этих (и многих других) экспериментов. Указывают, что вентральный зрительный тракт – вместилище зрительного осознания, по крайней мере, здесь находятся нейрональные корреляты содержания. Если объединить эти данные с независимыми данными, полученными у больных с нейропсихологическими расстройствами, потерявшими способность видеть зрительные образы (мы рассматривали эти данные в главе 4), мы наблюдаем убедительное единство в результатах, которое подтверждает, что для зрительного осознания того или иного содержания необходима активность одних и тех же анатомических областей вдоль вентрального зрительного тракта.
Вентральный зрительный тракт – это сложная совокупность участков коры головного мозга. Поэтому можно сказать, что активность в какой‑то ее части коррелирует со зрительным осознанием, не имея в виду совершенно точной локализации нейрональных механизмов сознания. До сих пор остается неясным, где и на каком участке вдоль вентрального тракта информация впервые всплывает в сознании. Почти все исследования указывают, что для зрительного осознания наиболее важны эволюционно более поздние области в височной доле. Но многие исследования также показывают, что активность в первичной зрительной коре (V1), куда сначала поступает зрительная информация извне, может отражать содержание зрительного осознания, а не физический стимул как таковой (Rees, 2007).
Роль области V1 в зрительном осознании до сих пор не выяснена. Есть данные, указывающие на ее участие, а также другие данные, указывающие на то, что она не является необходимой для зрительного осознания. Например, люди, у которых зона V1 оказалась повреждена вследствие травмы головного мозга и которые страдают кортикальной слепотой, все же могут представлять себе зрительные образы (Solms, 1997). Это доказывает, что зона V1 не является абсолютно необходимой для зрительного переживания. Даже если зона V1 так или иначе способствует зрительному осознанию, она делает это не одна. Вероятно, она взаимодействует с несколькими областями, расположенными дальше вдоль вентрального зрительного тракта, и зрительное содержание входит в сознание только после того, как пройдет весь этот сложный путь. (Эта идея подробнее обсуждается в разделе «Теория рекуррентной обработки» в главе 11).
|
|
Многие исследования с помощью томографии мозга показывают, что кроме вентрального зрительного тракта активируются другие области мозга, прежде всего в лобной и теменной коре. Пока не ясно, действительно эти области необходимы для зрительного осознания или они просто активируются при выполнении зрительных задач, поскольку такие задачи требуют участия когнитивных функций более высокого уровня: селективного нисходящего внимания и рабочей памяти. Лобные и теменные нейронные сети, в любом случае, важны для пространственного внимания и пространственного осознания, потому что повреждение этих областей связано с игнорированием стимулов в одной половине пространства (Rees, 2007).
Гипотеза о том, что лобные и теменные области не являются необходимыми для зрительного осознания, подтверждена экспериментами, показавшими, что когда селективное нисходящее внимание не является необходимым для выполнения зрительных задач, эти области не активируются, хотя информация о стимуле входит в зрительное осознание (Eriksson, Larsson, & Nyberg, 2008). Другие эксперименты показали, что активация лобных областей коррелирует не только со зрительным осознанием, но также и со слуховым осознанием (Eriksson, Larsson, Ahl strom, & Nyberg, 2007). Именно это мы обнаружим, если лобные области больше связаны со вниманием, независимым от модальностей, рефлексивным осознанием и рабочей памятью, а не с феноменальным осознанием определенной модальности.
Однако противоречия между результатами разных исследований и разными гипотезами относительно НКС и зрительного осознания отнюдь не разрешены. Некоторые из этих противоречий связаны с более глубокими теоретическими и философскими предположениями о природе сознания: существует ли такая вещь, как чистое феноменальное сознание? Отражается ли весь осознаваемый опыт в рефлексивном сознании? То есть зависит ли все осознаваемое зрительное содержание от селективного внимания, рабочей памяти и способности выражать его словами? Мы вернемся к этим вопросам, когда будем обсуждать другие теории сознания, где этот спор возникнет снова.
ЭЭГ и МЭГ исследования зрительного осознания
В экспериментах с ЭЭГ и МЭГ можно отследить во времени, когда информация от зрительного стимула входит в сознание. Сигналы ЭЭГ и МЭГ показывают, с временным масштабом в 1/1000 секунды, как развиваются электромагнитные реакции мозга. Сравнивая реакции мозга на увиденные (осознаваемые) стимулы со сходными стимулами, которые не были увидены, мы должны заметить, что в какой‑то момент мозг начинает по‑разному реагировать на стимулы, которые входят в зрительное осознание, и на стимулы, которые не осознаются. Когда это происходит? Когда и где мозг позволяет зрительному стимулу войти в сознание?
Основная идея экспериментов с ЭЭГ и МЭГ в сфере НКС нам уже знакома: сравнить, что происходит при наличие осознания стимулов и при отсутствии их осознания. Чтобы одни и те же стимулы иногда были невидимыми, а иногда видимыми, их часто демонстрируют на грани субъективного порога восприятия. Это значит, что время предъявления стимулов так невелико или их так трудно отличить от фона, что обычно испытуемый видит их лишь иногда и не видит в других случаях. При такой схеме эксперимента можно сравнить реакцию на увиденные и пропущенные стимулы при прочих равных (по крайней мере, очень близких к равным) условиях.
В подобных экспериментах одному и тому же испытуемому стимул предъявляют десятки или даже сотни раз, потому что реакция ЭЭГ на отдельный стимул настолько мала относительно всей остальной постоянной активности ЭЭГ, что ее почти невозможно различить. Нормальная постоянная ЭЭГ всегда содержит всевозможные виды спонтанной электрофизиологической активности мозга, не связанной ни с какими внешними стимулами. Слабые реакции ЭЭГ, каждый раз вызванные стимулом, усредняются, чтобы сформировать среднюю кривую реакций. Эта кривая нейтрализует всю активность ЭЭГ, не имеющую особого отношения к стимулу. Усредненную кривую реакций на стимул называют связанным с событием потенциалом (ССП), потому что она показывает только те электрические события в мозге, которые определенно связаны с интересующим нас событием, – в данном случае со зрительным стимулом.
Электрические реакции, связанные с событием, демонстрируют первые надежные различия между увиденными (осознаваемыми) и не увиденными (неосознаваемыми) стимулами. Эти различия возникают примерно на 150–200‑й миллисекунде после начала предъявления стимула и обычно достигают пика через 250–300 миллисекунд. В исследованиях ЭЭГ эту реакцию называют «негативностью зрительного осознания» (visual awareness negativity) (НЗО), потому что обычно на связанных с событием кривых потенциала мозга она выглядит как отрицательно направленная волна и возникает только при предъявлении стимулов, которые достигают сознания (Koivisto, Kainulainen. & Revonsuo, 2009; Wilenius & Revonsuo, 2007). Эта реакция сильнее всего обнаруживается в задней части головы, в местах, где электроды присоединены к затылочной, височной и задней теменной долям, – в областях скальпа, находящихся как раз над зрительной зоной коры головного мозга.
В исследовании МЭГ подобные реакции на увиденные объекты были локализованы в правой затылочной доле через 250–300 миллисекунд после начала предъявления стимула (Vanni, Revonsuo, Saarinen, & Hari, 1996). Нейрональная активность, которая выявляет НЗО, как полагают, является в мозге электрическим коррелятом феноменального зрительного осознания. Если эта гипотеза верна, мозгу нужно около 0,2–0,3 секунды на то, чтобы обработать информацию о стимуле в зрительной зоне коры головного мозга, – до того, как эта информация сможет войти в сознание.
Может показаться, что это очень быстро, но на самом деле для мозга это довольно медленная реакция. Зрительная информация входит в первичную зрительную кору уже в течение первых 20–30 миллисекунд и быстро обрабатывается во всех зрительных областях в несколько первых миллисекунд. Считается, что такая обработка происходит совершенно автоматически, за пределами сознания. Но при этом эта информация может управлять нашими быстрыми моторными реакциями на зрительные стимулы (в дело снова вступает лишенный сознания зрительно‑моторный зомби!).
Если вы когда‑либо рефлекторно отбивали теннисную подачу или бадминтонный воланчик, хотя даже не видели мяча или воланчика, по которому ударили, то ваша быстрая и успешная реакция, скорее всего, была результатом неосознаваемой быстрой обработки. Для осознания стимула, наоборот, нужен сложный синтез обратной связи между разными областями коры мозга, а это требует времени. Поэтому осознаваемое восприятие и основанные на нем действия обычно происходят гораздо медленнее, чем 0,1 секунды, которая требуется для первой быстрой реакции. (Подробнее о различиях между быстрой и медленной обработкой зрительных стимулов мы поговорим в обсуждении теории Ламме в главе 11).
За реакцией НЗО на ЭЭГ обычно следует другая волна, «поздняя позитивность» ( late positivity ) (ПП). Это медленная, широкая волна, возникающая примерно через 400–600 миллисекунд от начала действия стимула. Она очень похожа на другие позитивные волны типа P3 на ЭЭГ, которые, как полагают, отражают активность внимания и рабочей памяти. В экспериментах с осознанием поздняя позитивная волна считается коррелятом рефлексивного сознания: активности селективного внимания, классификации, называния и самоотчета об осознаваемом зрительном восприятии объекта (Koivislo, Kainulainen, & Revonsuo, 2009). Таким образом, исследования ЭЭГ также указывают на совокупность реакций, которые вполне можно интерпретировать как наличие феноменального сознания (на что указывает НЗО) и рефлексивного сознания (на что указывает ПП). Кроме того, поздняя позитивность не возникает, если стимул, достигающий феноменального сознания и генерирующий НЗО, не требует особого внимания, определенной реакции или сознательной рефлексии (Koivisto, Kainulainen, & Revonsuo, 2009).
Транскраниальная магнитная стимуляция (ТМС)
Недавно к арсеналу когнитивных неврологов добавился новый экспериментальный метод, дополняющий вышеупомянутые подходы. Метод транскраниальной магнитной стимуляции (ТМС) не похож на методы томографии мозга, ЭЭГ или МЭГ. Скорее, это способ локального вмешательства в нормальное функционирование мозга контролируемым образом. С помощью ТМС в определенные участки на поверхности коры мозга можно направить короткие магнитные импульсы. Они вызывают необычный взрыв нейроэлектрической активности, ненадолго прерывающий нормальную активность мозга. В экспериментах с ТМС можно наблюдать влияние таких вмешательств на поведение и сознание.
Когда с помощью этого устройства стимулируют зрительную зону коры мозга, можно наблюдать несколько эффектов. При низкой интенсивности магнитных импульсов (что создает лишь небольшой искусственный взрыв активности в зрительной зоне коры мозга), непосредственно стимулируя кору, например зрительные области V1 и V5, можно фактически создавать зрительное переживание . При стимулировании этих областей испытуемые сообщают о том, что видят мгновенные зрительные образы (их называют фосфенами ) – короткие вспышки света и зрительные образы. Фосфены бывают разной формы, иногда они имеют тот или иной цвет или движутся. По контрасту, если интенсивность импульсов ТМС высока, то подвергающаяся воздействию часть зрительной коры мозга, похоже, на время прекращает выполнять свои функции. Эта область становится временно слепой к зрительным стимулам (возникает «скотома», или слепая часть поля зрения), или, как минимум, в соответствующей части поля зрения снижается восприимчивость к стимулам.
Результаты исследований с использованием ТМС показывают, что когда возникают зрительные феномены, различные зрительные области взаимодействуют друг с другом. Например, когда возникают движущиеся фосфены, активизируются области V1 и V5. Если импульсы ТМС воздействуют на область V1 и в то же время на область V5, в попытке вызвать движущиеся фосфены, испытуемый не видит фосфенов. Таким образом, кажется, области V1 и V5 взаимодействуют друг с другом в том случае, когда возникают движущиеся зрительные феномены (Silvanto, Cowey, Lavie, & Walsh. 2005).
ТМС помогает установить, где расположены нейрональные механизмы зрительного осознания. Этот метод имеет преимущество перед другими, о которых мы говорили выше: с помощью ТМС можно выяснить не только то, какие области коррелируют с сознанием, но и то, какие из коррелирующих областей действительно важны и какого рода их активность необходима для зрительного осознания. Все остальные методы просто коррелятивны – они показывают, что осознание стимула и активность той или иной области имеют тенденцию возникать одновременно. Но такая корреляция не говорит о том, что именно делает эта область. С помощью ТМС можно помешать области выполнять свою задачу, а потом проверить, как отсутствие активности в этой области меняет сознание. Угнетая активность той или иной области и сравнивая полученные данные с данными ее нормальной работы, можно выяснить, по крайней мере косвенно, как нормальное функционирование этой области способствует осознанию. В этом смысле эксперименты с ТМС больше напоминают изучение кратковременных, небольших повреждений или временных нейропсихологических расстройств, чем создание функциональных карт при томографии мозга.
Выводы
В целом исследования с помощью функциональной томографии мозга и электромагнитного исследования мозга выявляют сходные данные относительно нейрональных коррелятов зрительного осознания. Когда зрительная информация включается в феноменальное сознание, активируются области коры мозга вдоль вентрального зрительного тракта. Эта активность быстро распространяется по всему пути, но чтобы зрительная информация достигла зрительного осознания, необходимо сложное взаимодействие, обмен обратной связью между разными областями коры мозга.
Поэтому чтобы информация включилась в сознание, нужно какое‑то время – как минимум около двух десятых секунды, и это отражается в хронологии того, как возникают первые надежные электрические реакции на сознание. Сразу же после того как информация впервые входит в феноменальное сознание, она направляется в рефлексивное сознание, особенно если эта информация необходима для выполнения текущей задачи, достижения цели или принятия решения. Этап рефлексивного сознания коррелирует с активацией лобно‑теменных областей мозга, которые, как известно, необходимы для внимания и рабочей памяти. Также известно, что с данными когнитивными механизмами коррелирует определенная волна связанных с событием потенциалов на ЭЭГ («поздняя позитивность»). Она указывает, что зрительная информация достигла рефлексивного сознания.
Такое представление о нейрональных коррелятах зрительного осознания поддерживают данные исследований больных с нейропсихологическими расстройствами, у которых отмечаются повреждения в вентральном зрительном тракте. Если вентральные области не только коррелируют со зрительным осознанием объектов, но и необходимы для его функционирования, то нарушения в этих областях должны приводить к нарушениям зрительного осознания, когда человек не видит объектов или каких‑то их особенностей.
Вот что при этом происходит: повреждение в области V4 приводит к ахроматопсии или исчезновению из зрительного осознания феноменальных цветов, повреждение латеральной затылочной коры (lateral occipital cortex) угнетает способность осознанно воспринимать целостные зрительные объекты (хотя действия под влиянием неосознаваемой зрительной информации все еще возможны), и т. д. (Подробнее см. более ранний раздел, посвященный нейропсихологии сознания, главы 4–6). Похожие эффекты можно также временно создавать у нормальных здоровых испытуемых с помощью ТМС зрительной зоны коры мозга.
Исследования в области нейрональных коррелятов сознания продолжаются, и в ведущих научных изданиях по когнитивной нейробиологии почти ежедневно публикуются новые результаты. Так что экспериментальный подход к исследованию НКС все ближе и ближе подводит нас к пониманию нейрональных механизмов сознания. Помогут ли все эти эмпирические данные окончательно решить философские проблемы, связанные с отношениями между сознанием и мозгом, пока неясно.
До сих пор нет никаких революционных данных или теорий, которые позволили бы хотя бы начать устранять «объяснительный разрыв». Даже если мы выясним где, когда и какая нейрональная активность участвует в создании зрительного переживания, поймем ли мы, как нейрональная активность возникает или как она создает субъективные зрительные феномены?
Вопросы для обсуждения
1. Придумайте самый лучший эксперимент НКС. Попробуйте представить себе, какой эксперимент мог бы выявить нейрональные корреляты сознания наиболее явным образом. Какой была бы задача испытуемого? Какие методы измерения вы бы использовали? Как результаты эксперимента помогли бы устранить объяснительный разрыв?
2. Если феноменальное сознание и рефлексивное сознание возникают почти одновременно, когда человек видит стимул, возможно ли измерить отдельно НКС для феноменального сознания и НКС для рефлексивного сознания? Какой эксперимент нужно провести, если мы хотим изучить НКС только для феноменального сознания?
III. Теории сознания
Введение: что такое теория сознания?
Науке, имеющей дело с сознанием, нужна теория, которая объясняет, что такое сознание и каково его место в мире. В этом разделе мы кратко обсудим некоторые из самых известных теорий сознания, выдвинутых в рамках науки о сознании. Эти теории разделены на два типа: философские (глава 10) и эмпирические (глава 11). Первые называются «философскими», потому что выдвинуты философами, а не учеными‑нейробиологами. Они основаны не на подробных эмпирических отчетах, а на общих метафизических решениях проблемы сознания. Эти теории радикально противоречат друг другу относительно фундаментальной природы сознания и его места в мире.
Их можно воспринимать как попытки найти метафизические точки отсчета или предпосылки эмпирической науки о сознании. В главе 11 мы рассмотрим современные эмпирические теории сознания, в первую очередь теории нейрональных основ сознания. Эти теории не занимаются метафизическими вопросами об истинной природе сознания, скорее они стремятся связать определенные феномены сознания с определенными когнитивными и нейрональными механизмами в мозге. При этом они также во многом противоречат друг другу. Однако, по крайней мере, есть несколько основных вопросов, по которым эти теории согласуются друг с другом. Такое единство между разными теориями позволяет надеяться, что когда‑нибудь будет создана единая, общая теория сознания.
Теория, как предполагается, должна объяснять факты, а «объяснение» помогает понять, что происходит в реальном мире. Теории помогают выяснить, какие феномены существуют в мире, из чего они состоят, как различные феномены взаимодействуют между собой, создавая причинно‑следственные связи, и, в целом, как устроен мир. Атомистическая теория материи объясняет, из каких элементарных единиц состоит вся физическая материя, клеточная теория в биологии объясняет, на что похожи микроскопические элементы биологических тканей и организмов, а эволюционная теория объясняет процесс, посредством которого возникли разные виды живых организмов на этой планете.
Теория сознания должна описывать и объяснять сознание: рассказывать нам, что такое сознание и как оно связано с другими явлениями в мире, в особенности с мозгом. Описать сознание означает дать его определение, ввести понятия, которые ясно и системно описывают основные особенности сознания, привести показательные примеры этого феномена и дифференцировать его от других феноменов, с которыми его можно легко перепутать. Объяснить сознание означает связать его с другими феноменами, описав механизмы и принципы, лежащие в его основе или несущие ответственность за его возникновение, и показав, как сознание взаимодействует с мозгом и как оно руководит нашим поведением.
Поэтому общее объяснение должно вести к глубокому пониманию природы , структуры , происхождения и функций того или иного феномена. Идеальная теория сознания должна предоставить объяснение, позволяющее понять сознание таким, каково оно есть, и определить его место в природе. По сути, сознание и есть то, что мы есть , – в конце концов, оно и создает переживание нашего субъективного, личного существования в этом мире. Теория сознания обязательно также скажет нам, каково наше место во Вселенной: мы – фрагменты вечной нематериальной субстанции души, или хрупкие, временные совокупности биоэлектрических полей в мозге, или мы все время ошибались и сознание на самом деле нечто совсем другое, не соответствующее нашим интуитивным догадкам и традиционным объяснениям.
Общая теория сознания должна также во всех подробностях объяснять или предсказывать, что происходит с сознанием в исключительных обстоятельствах, например при различных травмах мозга или во время сна, анестезии, внетелесного и предсмертного переживания. Существование и особенности такого переживания должны естественно и логично следовать из Единой теории сознания. Я уверен, что нам всем было бы очень любопытно увидеть такую теорию!
В современной литературе представлено множество теорий сознания. К сожалению, ни одну из них пока нельзя назвать Единой теорией. Наличие множества разнообразных теорий сознания – это и хорошо, и плохо. Хорошо в том смысле, что разные теории исследуют разные подходы к объяснению сознания и поэтому помогают находить самые плодотворные подходы, ведущие к наиболее правдоподобным объяснениям. А плохо, потому что все эти теории настолько разнообразны, что даже неясно, описывают ли они все одно и то же явление , когда используют слово «сознание». Кроме того, почти все теории концентрируется на объяснении нескольких избранных аспектов осознаваемого переживания. Остается неясным, можно ли применить их также и к другим явлениям как к целому, на общем уровне.
В любом случае, окончательная цель науки о сознании, конечно, заключается не в получении миллиона разных теорий сознания, несовместимых друг с другом, а скорее в создании Единой теории сознания, описывающей и объясняющей все возможные типы феноменов сознания раз и навсегда. Но сейчас, на ранней стадии развития науки о сознании, мы даже не знаем, в каком направлении нужно искать общую теорию сознания. Философы ищут объяснения сознания повсюду. Давайте же посмотрим, что они предлагают. Возможно, это и есть правильный путь.
Глава 10
Философские теории сознания
|
|
|

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Таксономические единицы (категории) растений: Каждая система классификации состоит из определённых соподчиненных друг другу...

Наброски и зарисовки растений, плодов, цветов: Освоить конструктивное построение структуры дерева через зарисовки отдельных деревьев, группы деревьев...

Архитектура электронного правительства: Единая архитектура – это методологический подход при создании системы управления государства, который строится...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!