Строение клетки
Строение прокариотических и эукариотических клеток
Основными структурными компонентами клеток являются плазматическая мембрана, цитоплазма и наследственный аппарат. В зависимости от особенностей организации различают два основных типа клеток: прокариотические и эукариотические. Главным отличием прокариотических клеток от эукариотических является организация их наследственного аппарата: у прокариот он находится непосредственно в цитоплазме (эта область цитоплазмы называется нуклеоидом) и не отделен от нее мембранными структурами, тогда как у эукариот бульшая часть ДНК сосредоточена в ядре, окруженном двойной мембраной. Кроме того, генетическая информация прокариотических клеток, находящаяся в нуклеоиде, записана в кольцевой молекуле ДНК, а у эукариот молекулы ДНК незамкнутые.
В отличие от эукариот, цитоплазма прокариотических клеток содержит также небольшое количество органоидов, тогда как для эукариотических характерно значительное разнообразие этих структур.
Цитоплазма
Цитоплазма — это внутреннее содержимое клетки. В нее погружены все органоиды клетки, ядро и разнообразные продукты жизнедеятельности.
Цитоплазма связывает все части клетки между собой, в ней протекают многочисленные реакции обмена веществ. Цитоплазма отделяется от окружающей среды и делится на отсеки мембранами, то есть клеткам присуще мембранное строение. Она может находиться в двух состояниях — золя и геля. Золь — это полужидкое, киселеобразное состояние цитоплазмы, при котором процессы жизнедеятельности протекают наиболее интенсивно, а гель — более плотное, студнеобразное состояние, затрудняющее протекание химических реакций и транспорт веществ.
Жидкая часть цитоплазмы без органоидов называется гиалоплазмой. Гиалоплазма, или цитозоль, представляет собой коллоидный раствор, в котором находится своеобразная взвесь достаточно крупных частиц, например белков, окруженных диполями молекул воды. Осаждения этой взвеси не происходит вследствие того, что они имеют одинаковый заряд и отталкиваются друг от друга.
Органоиды
Органоиды — это постоянные компоненты клетки, выполняющие определенные функции.
В зависимости от особенностей строения их делят на мембранные и немембранные. Мембранные органоиды, в свою очередь, относят к одномембранным (эндоплазматическая сеть, комплекс Гольджи и лизосомы) или двумембранным (митохондрии, пластиды и ядро). Немембранными органоидами являются рибосомы, микротрубочки, микрофиламенты и клеточный центр. Прокариотам из перечисленных органоидов присущи только рибосомы.
Строение и функции ядра. Ядро — крупный двумембранный органоид, лежащий в центре клетки или на ее периферии. Размеры ядра могут колебаться в пределах 3–35 мкм. Форма ядра чаще сферическая или эллипсоидная, однако имеются также палочковидные, веретеновидные, бобовидные, лопастные и даже сегментированные ядра. Некоторые исследователи считают, что форма ядра соответствует форме самой клетки.
Большинство клеток имеет одно ядро, но, например, в клетках печени и сердца их может быть два, а в ряде нейронов — до 15. Волокна скелетных мышц содержат обычно много ядер, однако они не являются клетками в полном смысле этого слова, поскольку образуются в результате слияния нескольких клеток.
Ядро окружено ядерной оболочкой, а его внутреннее пространство заполнено ядерным соком, или нуклеоплазмой (кариоплазмой), в которую погружены хроматин и ядрышко. Ядро выполняет такие важнейшие функции, как хранение и передача наследственной информации, а также контроль жизнедеятельности клетки.

Роль ядра в передаче наследственной информации была убедительно доказана в экспериментах с зеленой водорослью ацетабулярией. В единственной гигантской клетке, достигающей в длину 5 см, различают шляпку, ножку и ризоид. При этом она содержит только одно ядро, расположенное в ризоиде. В 1930-е годы И. Хеммерлинг пересадил ядро одного вида ацетабулярии с зеленой окраской в ризоид другого вида, с коричневой окраской, у которого ядро было удалено. Через некоторое время у растения с пересаженным ядром выросла новая шляпка, как у водоросли- донора ядра. В то же время отделенные от ризоида шляпка или ножка, не содержащие ядра, через некоторое время погибали.

Ядерная оболочка образована двумя мембранами — наружной и внутренней, между которыми есть пространство. Межмембранное пространство сообщается с полостью шероховатой эндоплазматической сети, а наружная мембрана ядра может нести рибосомы. Ядерная оболочка пронизана многочисленными порами, окантованными специальными белками. Через поры происходит транспорт веществ: в ядро попадают необходимые белки (в т. ч. ферменты), ионы, нуклеотиды и другие вещества, и покидают его молекулы РНК, отработанные белки, субъ единицы рибосом. Таким образом, функциями ядерной оболочки являются отделение содержимого ядра от цитоплазмы, а также регуляция обмена веществ между ядром и цитоплазмой.
Нуклеоплазмой называют содержимое ядра, в которое погружены хроматин и ядрышко. Она представляет собой коллоидный раствор, по химическому составу напоминающий цитоплазму. Ферменты нуклеоплазмы катализируют обмен аминокислот, нуклеотидов, белков и др. Нуклеоплазма связана с гиалоплазмой через ядерные поры. Функции нуклеоплазмы, как и гиалоплазмы, состоят в обеспечении взаимосвязи всех структурных компонентов ядра и осуществлении ряда ферментных реакций.
Хроматином называют совокупность тонких нитей и гранул, погруженных в нуклеоплазму. Выявить его можно только при окрашивании, так как коэффициенты преломления хроматина и нуклеоплазмы приблизительно одинаковы. Нитчатый компонент хроматина называют эухроматином, а гранулярный — гетерохроматином. Эухроматин слабо уплотнен, поскольку с него считывается наследственная информация, тогда как более спирализованный гетерохроматин является генетически неактивным.
Хроматин представляет собой структурное видоизменение хромосом в неделящемся ядре. Таким образом, хромосомы постоянно присутствуют в ядре, изменяется лишь их состояние в зависимости от функции, которую ядро выполняет в данный момент.
В состав хроматина в основном входят белки-нуклеопротеины (дезоксирибонуклеопротеины и рибонуклеопротеины), а также ферменты, важнейшие из которых связаны с синтезом нуклеиновых кислот, и некоторые другие вещества.
Функции хроматина состоят, во-первых, в синтезе специфических для данного организма нуклеиновых кислот, которые направляют синтез специфических белков, во-вторых, в передаче наследственных свойств от материнской клетки дочерним, для чего хроматиновые нити в процессе деления упаковываются в хромосомы.
Ядрышко — сферическое, хорошо заметное под микроскопом тельце диаметром 1–3 мкм. Оно формируется на участках хроматина, в которых закодирована информация о структуре рРНК и белках рибосом. Ядрышко в ядре часто одно, однако в тех клетках, где происходят интенсивные процессы жизнедеятельности, ядрышек может быть два и более. Функции ядрышек — синтез рРНК и сборка субъединиц рибосом путем объединения рРНК с белками, поступающими из цитоплазмы.
Митохондрии — двумембранные органоиды округлой, овальной или палочковидной формы, хотя встречаются и спиралевидные (в сперматозоидах). Диаметр митохондрий составляет до 1 мкм, а длина — до 7 мкм. Пространство внутри митохондрий заполнено матриксом. Матрикс — это основное вещество митохондрий. В него погружены кольцевая молекула ДНК и рибосомы. Наружная мембрана митохондрий гладкая, она непроницаема для многих веществ. Внутренняя мембрана имеет выросты — кристы, увеличивающие площадь поверхности мембран для протекания химических реакций. На поверхности мембраны расположены многочисленные белковые комплексы, составляющие так называемую дыхательную цепь, а также грибовидные ферменты АТФ-синтетазы. В митохондриях протекает аэробный этап дыхания, в ходе которого происходит синтез АТФ.

Пластиды — крупные двумембранные органоиды, характерные только для растительных клеток. Внутреннее пространство пластид заполнено стромой, или матриксом. В строме находится более или менее развитая система мембранных пузырьков — тилакоидов, которые собраны в стопки — граны, а также собственная кольцевая молекула ДНК и рибосомы. Различают четыре основных типа пластид: хлоропласты, хромопласты, лейкопласты и пропластиды.
Хлоропласты — это зеленые пластиды диаметром 3–10 мкм, хорошо различимые под микроскопом. Они содержатся только в зеленых частях растений — листьях, молодых стеблях, цветках и плодах. Хлоропласты в основном имеют овальную или эллипсоидную формы, но могут быть также чашевидными, спиралевидными и даже лопастными. Количество хлоропластов в клетке в среднем составляет от 10 до 100 штук. Однако, например, у некоторых водорослей он может быть один, иметь значительные размеры и сложную форму — тогда его называют хроматофором. В других случаях количество хлоропластов может достигать нескольких сотен, при этом их размеры невелики. Окраска хлоропластов обусловлена основным пигментом фотосинтеза — хлорофиллом, хотя в них содержатся и дополнительные пигменты — каротиноиды. Каротиноиды становятся заметными только осенью, когда хлорофилл в стареющих листьях разрушается. Основной функцией хлоропластов является фотосинтез. Световые реакции фотосинтеза протекают на мембранах тилакоидов, на которых закреплены молекулы хлорофилла, а темновые реакции — в строме, где содержатся многочисленные ферменты.

Хромопласты — это желтые, оранжевые и красные пластиды, содержащие пигменты каротиноиды. Форма хромопластов может также существенно варьировать: они бывают трубчатыми, сферическими, кристаллическими и др. Хромопласты придают окраску цветкам и плодам растений, привлекая опылителей и распространителей семян и плодов.
Лейкопласты — это белые или бесцветные пластиды в основном округлой или овальной формы. Они распространены в нефотосинтезирующих частях растений, например в кожице листа, клубнях картофеля и т. д. В них откладываются в запас питательные вещества, чаще всего крахмал, но у некоторых растений это могут быть белки или масло.
Пластиды образуются в растительных клетках из пропластид, которые имеются уже в клетках образовательной ткани и представляют собой небольшие двумембранные тельца. На ранних этапах развития разные виды пластид способны превращаться друг в друга: при попадании на свет лейкопласты клубня картофеля и хромопласты корнеплода моркови зеленеют.
Пластиды и митохондрии называют полуавтономными органоидами клетки, так как они имеют собственные молекулы ДНК и рибосомы, осуществляют синтез белка и делятся независимо от деления клеток. Эти особенности объясняются происхождением от одноклеточных прокариотических организмов. Однако «самостоятельность» митохондрий и пластид является ограниченной, так как их ДНК содержит слишком мало генов для свободного существования, остальная же информация закодирована в хромосомах ядра, что позволяет ему контролировать данные органоиды.
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭР), — это одномембранный органоид, представляющий собой сеть мембранных полостей и канальцев, занимающих до 30 % содержимого цитоплазмы. Диаметр канальцев ЭПС составляет около 25–30 нм. Различают два вида ЭПС — шероховатую и гладкую. Шероховатая ЭПС несет рибосомы, на ней происходит синтез белков. Гладкая ЭПС лишена рибосом. Ее функция — синтез липидов и углеводов, а также транспорт, запасание и обезвреживание токсических веществ. Она особенно развита в тех клетках, где происходят интенсивные процессы обмена веществ, например в клетках печени — гепатоцитах — и волокнах скелетных мышц. Вещества, синтезированные в ЭПС, транспортируются в аппарат Гольджи. В ЭПС происходит также сборка мембран клетки, однако их формирование завершается в аппарате Гольджи.
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид, образованный системой плоских цистерн, канальцев и отшнуровывающихся от них пузырьков. Структурной единицей аппарата Гольджи является диктиосома — стопка цистерн, на один полюс которой приходят вещества из ЭПС, а с противоположного полюса, подвергшись определенным превращениям, они упаковываются в пузырьки и направляются в другие части клетки. Диаметр цистерн — порядка 2 мкм, а мелких пузырьков — около 20–30 мкм. Основные функции комплекса Гольджи — синтез некоторых веществ и модификация (изменение) белков, липидов и углеводов, поступающих из ЭПС, окончательное формирование мембран, а также транспорт веществ по клетке, обновление ее структур и образование лизосом. Свое название аппарат Гольджи получил в честь итальянского ученого Камилло Гольджи, впервые обнаружившего данный органоид (1898).

Лизосомы — небольшие одномембранные органоиды до 1 мкм в диаметре, в которых содержатся гидролитические ферменты, участвующие во внутриклеточном пищеварении. Мембраны лизосом слабопроницаемы для этих ферментов, поэтому выполнение лизосомами своих функций происходит очень точно и адресно. Так, они принимают активное участие в процессе фагоцитоза, образуя пищеварительные вакуоли, а в случае голодания или повреждения определенных частей клетки переваривают их, не затрагивая иных. Недавно была открыта роль лизосом в процессах клеточной гибели.
Вакуоль — это полость в цитоплазме растительных и животных клеток, ограниченная мембраной и заполненная жидкостью. В клетках простейших обнаруживаются пищеварительные и сократительные вакуоли. Первые принимают участие в процессе фагоцитоза, так как в них происходит расщепление питательных веществ. Вторые обеспечивают поддержание водно-солевого баланса за счет осморегуляции. У многоклеточных животных в основном встречаются пищеварительные вакуоли.
В растительных клетках вакуоли присутствуют всегда, они окружены специальной мембраной и заполнены клеточным соком. Мембрана, окружающая вакуоль, по химическому составу, строению и выполняемым функциям близка к плазматической мембране. Клеточный сок представляет собой водный раствор различных неорганических и органических веществ, в том числе минеральных солей, органических кислот, углеводов, белков, гликозидов, алкалоидов и др. Вакуоль может занимать до 90 % объема клетки и оттеснять ядро на периферию. Эта часть клетки выполняет запасающую, выделительную, осмотическую, защитную, лизосомную и другие функции, поскольку в ней накапливаются питательные вещества и отходы жизнедеятельности, она обеспечивает поступление воды и поддержание формы и объема клетки, а также содержит ферменты расщепления многих компонентов клетки. К тому же биологически активные вещества вакуолей способны препятствовать поеданию этих растений многими животными. У ряда растений за счет разбухания вакуолей происходит рост клетки растяжением.
Вакуоли имеются также и в клетках некоторых грибов и бактерий, однако у грибов они выполняют только функцию осморегуляции, а у цианобактерий поддерживают плавучесть и участвуют в процессах усвоения азота из воздуха.
Рибосомы — небольшие немембранные органоиды диаметром 15–20 мкм, состоящие из двух субъединиц — большой и малой. Субъединицы рибосом эукариот собираются в ядрышке, а затем транспортируются в цитоплазму. Рибосомы прокариот, митохондрий и пластид меньше по величине, чем рибосомы эукариот. В состав субъединиц рибосом входят рРНК и белки.

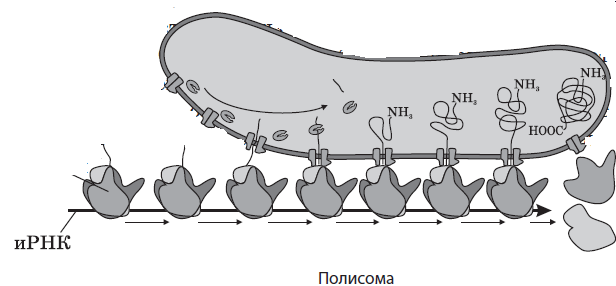
Количество рибосом в клетке может достигать нескольких десятков миллионов: в цитоплазме, митохондриях и пластидах они находятся в свободном состоянии, а на шероховатой ЭПС — в связанном. Они принимают участие в синтезе белка, в частности, осуществляют процесс трансляции — биосинтеза полипептидной цепи на молекуле иРНК. На свободных рибосомах синтезируются белки гиалоплазмы, митохондрий, пластид и собственные белки рибосом, тогда как на прикрепленных к шероховатой ЭПС рибосомах осуществляется трансляция белков для выведения из клеток, сборки мембран, образования лизосом и вакуолей.
Рибосомы могут находиться в гиалоплазме поодиночке или собираться в группы при одновременном синтезе на одной иРНК сразу нескольких полипептидных цепей. Такие группы рибосом называются полирибосомами, или полисомами.
Микротрубочки — это цилиндрические полые немембранные органоиды, которые пронизывают всю цитоплазму клетки. Их диаметр составляет около 25 нм, толщина стенки — 6–8 нм. Они образованы многочисленными молекулами белка тубулина, которые сначала формируют 13 нитей, напоминающих бусы, а затем собираются в микротрубочку. Микротрубочки образуют цитоплазматическую сеть, которая придает клетке форму и объем, связывают плазматическую мембрану с другими частями клетки, обеспечивают транспорт веществ по клетке, принимают участие в движении клетки и внутриклеточных компонентов, а также в делении генетического материала. Они входят в состав клеточного центра и органоидов движения — жгутиков и ресничек.
Микрофиламенты, или микронити, также являются немембранными органоидами, однако они имеют нитевидную форму и образованы не тубулином, а актином. Они принимают участие в процессах мембранного транспорта, межклеточном узнавании, делении цитоплазмы клетки и в ее движении. В мышечных клетках взаимодействие актиновых микрофиламентов с миозиновыми нитями обеспечивает сокращение.
Микротрубочки и микрофиламенты образуют внутренний скелет клетки — цитоскелет. Он представляет собой сложную сеть волокон, обеспечивающих механическую опору для плазматической мембраны, определяет форму клетки, расположение клеточных органоидов и их перемещение в процессе деления клетки.

Клеточный центр — немембранный органоид, располагающийся в животных клетках вблизи ядра; в растительных клетках он отсутствует. Его длина составляет около 0.2–0.3 мкм, а диаметр — 0.1–0.15 мкм. Клеточный центр образован двумя центриолями, лежащими во взаимно перпендикулярных плоскостях, и лучистой сферой из микротрубочек. Каждая центриоль образована девятью группами микротрубочек, собранных по три, т. е. триплетами. Клеточный центр принимает участие в процессах сборки микротрубочек, делении наследственного материала клетки, а также в образовании жгутиков и ресничек.

Органоиды движения. Жгутики и реснички представляют собой выросты клетки, покрытые плазмалеммой. Основу этих органоидов составляют девять пар микротрубочек, расположенных по периферии, и две свободные микротрубочки в центре. Микротрубочки связаны между собой различными белками, обеспечивающими их согласованное отклонение от оси — колебание. Колебания энергозависимы, то есть на этот процесс тратится энергия макроэргических связей АТФ. Восстановление утраченных жгутиков и ресничек является функцией базальных телец, или кинетосом, расположенных в их основании.
Длина ресничек составляет около 10–15 нм, а жгутиков — 20–50 мкм. За счет строго направленных движений жгутиков и ресничек осуществляется не только движение одноклеточных животных, сперматозоидов и др., но и происходит очистка дыхательных путей, продвижение яйцеклетки по маточным трубам, поскольку все эти части организма человека выстланы реснитчатым эпителием.

Включения
Включения — это непостоянные компоненты клетки, которые образуются и исчезают в процессе ее жизнедеятельности. К ним относят как запасные вещества, например, зерна крахмала или белка в растительных клетках, гранулы гликогена в клетках животных и грибов, волютина у бактерий, капли жира во всех типах клеток, так и отходы жизнедеятельности, в частности, непереваренные в результате фагоцитоза остатки пищи, образующие так называемые остаточные тельца.

Обмен веществ и превращения энергии — свойства живых организмов. Энергетический и пластический обмен, их взаимосвязь. Стадии энергетического обмена. Брожение и дыхание. Фотосинтез, его значение, космическая роль. Фазы фотосинтеза. Световые и темновые реакции фотосинтеза, их взаимосвязь. Хемосинтез. Роль хемосинтезирующих бактерий на Земле
Дыхание
Механизм клеточного дыхания схематически изображен на рис.

Аэробное дыхание происходит в митохондриях, при этом пировиноградная кислота сначала утрачивает один атом углерода, что сопровождается синтезом одного восстановительного эквивалента НАДН+Н+ и молекулы ацетилкофермента А (ацетил-КоА):
С3Н4О3+НАД+Н~КоА→СН3СО~КоА+НАДН+Н++СО2↑.
Ацетил-КоА в матриксе митохондрий вовлекается в цепь химических реакций, совокупность которых называется циклом Кребса (циклом трикарбоновых кислот, циклом лимонной кислоты). В ходе этих превращений образуется две молекулы АТФ, ацетил-КоА полностью окисляется до углекислого газа, а его ионы водорода и электроны присоединяются к переносчикам водорода НАДН+Н+ и ФАДН2. Переносчики транспортируют протоны водорода и электроны к внутренним мембранам митохондрий, образующим кристы. При помощи белков-переносчиков протоны водорода нагнетаются в межмембранное пространство, а электроны передаются по так называемой дыхательной цепи ферментов, расположенной на внутренней мембране митохондрий, и сбрасываются на атомы кислорода:
O2+2e−→O2−.
Следует отметить, что некоторые белки дыхательной цепи содержат железо и серу.
Из межмембранного пространства протоны водорода транспортируются обратно в матрикс митохондрий с помощью специальных ферментов — АТФ-синтаз, а выделяющаяся при этом энергия расходуется на синтез 34 молекул АТФ из каждой молекулы глюкозы. Этот процесс называется окислительным фосфорилированием. В матриксе митохондрий протоны водорода реагируют с радикалами кислорода с образованием воды:
4H++O2−→2H2O.
Совокупность реакций кислородного дыхания может быть выражена следующим образом:
2С3Н4О3+6О2+36Н3РО4+36АДФ→6СО2↑+38Н2О+36АТФ.
Суммарное уравнение дыхания выглядит таким образом:
С6Н12О6+6О2+38Н3РО4+38АДФ→6СО2↑+40Н2О+38АТФ.
Брожение
В отсутствие кислорода или при его недостатке происходит брожение. Брожение является эволюционно более ранним способом получения энергии, чем дыхание, однако оно энергетически менее выгодно, поскольку в результате брожения образуются органические вещества, все еще богатые энергией. Различают несколько основных видов брожения: молочнокислое, спиртовое, уксуснокислое и др. Так, в скелетных мышцах в отсутствие кислорода в ходе брожения пировиноградная кислота восстанавливается до молочной кислоты, при этом образовавшиеся ранее восстановительные эквиваленты расходуются, и остаются всего две молекулы АТФ:
2С3Н4О3+2НАДН+Н+→2С3Н6О3+2НАД.
При брожении с помощью дрожжевых грибов пировиноградная кислота в присутствии кислорода превращается в этиловый спирт и оксид углерода (IV):
С3Н4О3+НАДН+Н+→С2Н5ОН+СО2↑+НАД+.
При брожении с помощью микроорганизмов из пировиноградной кислоты могут образоваться также уксусная, масляная, муравьиная кислоты и др.
АТФ, полученная в результате энергетического обмена, расходуется в клетке на различные виды работы: химическую, осмотическую, электрическую, механическую и регуляторную. Химическая работа заключается в биосинтезе белков, липидов, углеводов, нуклеиновых кислот и других жизненно важных соединений. К осмотической работе относят процессы поглощения клеткой и выведения из нее веществ, которые во внеклеточном пространстве находятся в концентрациях, больших, чем в самой клетке. Электрическая работа тесно взаимосвязана с осмотической, поскольку именно в результате перемещения заряженных частиц через мембраны формируется заряд мембраны и приобретаются свойства возбудимости и проводимости. Механическая работа сопряжена с движением веществ и структур внутри клетки, а также клетки в целом. К регуляторной работе относят все процессы, направленные на координацию процессов в клетке.
Клетка — генетическая единица живого. Хромосомы, их строение (форма и размеры) и функции. Число хромосом и их видовое постоянство. Соматические и половые клетки. Жизненный цикл клетки: интерфаза и митоз. Митоз — деление соматических клеток. Мейоз. Фазы митоза и мейоза. Развитие половых клеток у растений и животных. Деление клетки — основа роста, развития и размножения организмов. Роль мейоза и митоза
Мейоз. Фазы мейоза
Мейоз — это способ непрямого деления первичных половых клеток (2n2с), в результате которого образуются гаплоидные клетки (1n1с), чаще всего половые.
В отличие от митоза, мейоз состоит из двух последовательных делений клетки, каждому из которых предшествует интерфаза. Первое деление мейоза (мейоз I) называется редукционным, так как при этом количество хромосом уменьшается вдвое, а второе деление (мейоз II) — эквационным, так как в его процессе количество хромосом сохраняется.

Интерфаза I протекает подобно интерфазе митоза. Мейоз I делится на четыре фазы: профазу I, метафазу I, анафазу I и телофазу I. В профазе I происходят два важнейших процесса — конъюгация и кроссинговер. Конъюгация — это процесс слияния гомологичных (парных) хромосом по всей длине. Образовавшиеся в процессе конъюгации пары хромосом сохраняются до конца метафазы I.
Кроссинговер — взаимный обмен гомологичными участками гомологичных хромосом. В результате кроссинговера хромосомы, полученные организмом от обоих родителей, приобретают новые комбинации генов, что обусловливает появление генетически разнообразного потомства. В конце профазы I, как и в профазе митоза, исчезает ядрышко, центриоли расходятся к полюсам клетки, а ядерная оболочка распадается.

В метафазе I пары хромосом выстраиваются по экватору клетки, к их центромерам прикреп ляются микротрубочки веретена деления.
В анафазе I к полюсам расходятся целые гомологичные хромосомы, состоящие из двух хроматид.
В телофазе I вокруг скоплений хромосом у полюсов клетки образуются ядерные оболочки, формируются ядрышки.
Цитокинез I обеспечивает разделение цитоплазм дочерних клеток.
Образовавшиеся в результате мейоза I дочерние клетки (1n2c) генетически разнородны, поскольку их хромосомы, случайным образом разошедшиеся к полюсам клетки, содержат неодинаковые гены.
Строение клетки
Строение прокариотических и эукариотических клеток
Основными структурными компонентами клеток являются плазматическая мембрана, цитоплазма и наследственный аппарат. В зависимости от особенностей организации различают два основных типа клеток: прокариотические и эукариотические. Главным отличием прокариотических клеток от эукариотических является организация их наследственного аппарата: у прокариот он находится непосредственно в цитоплазме (эта область цитоплазмы называется нуклеоидом) и не отделен от нее мембранными структурами, тогда как у эукариот бульшая часть ДНК сосредоточена в ядре, окруженном двойной мембраной. Кроме того, генетическая информация прокариотических клеток, находящаяся в нуклеоиде, записана в кольцевой молекуле ДНК, а у эукариот молекулы ДНК незамкнутые.
В отличие от эукариот, цитоплазма прокариотических клеток содержит также небольшое количество органоидов, тогда как для эукариотических характерно значительное разнообразие этих структур.





