Жгутики
Капсула
Включения Рибосомы
Нуклеоид
Плазмида
Мезосома
Клеточная стенка
Цитоплазматическая мембрана
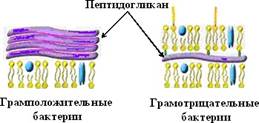
Бактериальная оболочка состоит из клеточной стенки и располагающейся под ней цитоплазматической мембраны. Клеточная стенка - это ригидная структу- ра, которая придает бактериальной клетке определенную форму. Она защищает внутреннее содержимое клетки от вредных воздействий внешней среды, участвует в процессах деления клетки и транспорта метаболитов. На поверхности клеточной стенки располагаются рецепторы для бактериофагов, бактериоцинов, антибиотиков и других химических веществ. По строению клеточной стенки различают фирмикутные бактерии (грамположительные, толстостенные), грациликутные бактерии (грамотрицательные, тонкостенные) и бактерии, не имеющие клеточной стенки (микоплазмы). Подразделение бактерий на грамположительные (грампозитивные) и грамотрицательные (грамнегативные) основано на разном восприятии красителей при окраске по методу, предложенному датским бактериологом Г. К. Грамом.
Клеточная стенка грамположительных бактерий представляет собой гомогенный слой. Она состоит из многослойного пептидогликана (муреина), пронизанного молекулами тейхоевой и липотейхоевой кислот.
Пептидогликан клеточной стенки образован параллельно расположенными молекулами гликана, состоящего из остатков N-ацетилглюкозамина и N- ацетилмурамовой кислоты, соединенных вдоль гликозидной связью.
Тейхоевые кислоты представляют собой цепи из остатков глицерола и рибитола, соединенных фосфатными мостиками. Пептидогликан и тейхоевые кислоты в конечном итоге формируют так называемый муреиновый мешок,покрывающий клетку снаружи. Тейхоевые кислоты позволяют муреиновому мешку растягиваться и сжиматься, действуя наподобие пружин. Тейхоевые кислоты выполняют антигенную и адгезивную функции грамположительных бактерий. Пептидогликан грамположительных бактерий плотно прилегает к цитоплазматичекой мембране. Клеточная стенка грамположительных бактерий содержит также небольшое количество полисахаридов, белков и липидов. В клеточной стенке имеются поры диаметром 1-6 нм, через которые внутрь клетки проникают различные вещества.
При окраске по Граму толстый слой пептидогликана грамположительных бактерий удерживает генциановый фиолетовый в комплексе с йодом. Последующая обработка препарата спиртом вызывает суживание пор в пептидогликане и тем самым усиливает задержку фиолетового красителя в клеточной стенке. Заключительная окраска препарата фуксином не изменяет первоначальной окраски клеток. Грамположительные бактерии окрашиваются в сине-фиолетовый цвет.
Клеточная стенка грамотрицательных бактерий выделяют внешнюю (наружную) мембрану (НМ) и тонкий пептидогликановый слой или муреиновый мешок (рисунок 37).
Внешняя мембрана грамотрицательных бактерий представляет собой фосфолипидный бислой, содержащий белки и липополисахарид. Липополисахарид (ЛПС) состоит из трех составных частей:
- липид А - специфический гликолипид, встроенной в фосфолипидный бислой, закрепляющий молекулу ЛПС во внешней мембране и придающий липополисахариду токсические свойства;
- ядро - центральная (стержневая, коровая) область (лат. core - ядро) полисахаридной природы;
- боковая О-цепь, образованная повторяющимися олигосахаридами (О- антиген).
Боковая О-цепь
Ядро
Липид А
С помощью мембранного липопротеина внешняя мембрана связана с подлежащим слоем пептидогликана. Пептидогликан грамотрицательных бактерий является однослойным (толщина – 2-3 нм) и не содержит тейхоевых кислот. Под слоем пептидогликана располагается цитоплазматическая мембрана. Между внешней мембраной, слоем пептидогликана и цитоплазматической мембраной имеется полость, называемая периплазматическим пространством (периплазмой). Это пространство заполнено гелем, содержащим транспортные белки и ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета- лактамазы). В периплазматическом геле располагается муреиновый (пептидогликановый) мешок.
Белки наружной мембраны включают порины, трансмембранные белки и белки, участвующие в формировании поверхностных структур (пилей, жгутиков). Порины образуют каналы для проникновения воды и мелких молекул. Трансмембранные белки обеспечивают связь с муреиновым мешком.
При окраске по Граму тонкий слой пептидогликана грамотрицательных бактерий под воздействием этилового спирта утрачивает комплекс генцианового фиолетового и йода. При последующей обработке фуксином или сафранином клетки приобретают красный цвет.
Рисунок 40 - Клеточные стенки грамположительных и грамотрицательных
бактерий.
К грамположительным бактериям относятся все кокки (за исключением гонококков и менингококков), спорообразующие палочки (бациллы и клостридии), микобактерии, коринебактерии, листерии. К грамотрицательным бактериям относятся гонококки и менингококки, не образующие спор палочки, извитые бактерии.
Клеточная стенка бактерий выполняет следующие функции:
- предохраняет клетку от вредных воздействий окружающей среды;
- обеспечивает постоянство формы клетки;
- сообщает бактериальной клетке антигенные свойства;
- регулирует рост и деление клетки;
- участвует в поступлении внутрь клетки некоторых молекул;
- обеспечивает тинкториальные свойства бактерий (отношение к красителям).
Под воздействием некоторых веществ (лизоцим, пенициллин, гуморальные факторы организма) нарушается синтез компонентов клеточной стенки бактерий. В таких случаях бактерии полностью или частично лишаются клеточной стенки, образуя шаровидные формы. Такие формы имеют размеры, превышающие исходные клетки в несколько раз. Бактерии, полностью лишенные клеточной стенки, называются протопластами, а бактерии, частично сохранившие клеточную стенку, называются сферопластами.
Образование протопластов характерно для грамположительных бактерий. Протопластообразование сопровождается утратой толстой пептидогликановой клеточной стенки. Протопласты содержат только цитоплазматическую мембрану (рисунок 42). Для их поддержания требуется изотоническая среда. Они устойчивы к антибиотикам и бактериофагам.
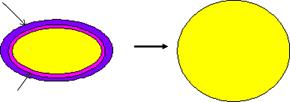 Клеточная стенка
Клеточная стенка
Цитоплазматическая мембрана
Сферопласты образуются грамотрицательными бактериями. Образование сферопластов сопровождается утратой внешней мембраны клеточной стенки. Но сферопласты наряду с цитоплазматической мембраной содержат тонкий слой пептидогликана (рисунок 43).
Пептидогликан
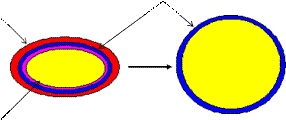 Клеточная стенка
Клеточная стенка
Цитоплазматическая мембрана
Рисунок 43 – Образование сферопластов у бактерий.
Для поддержания сферопластов также требуется среда с повышенным осмотическим давлением. Сферопласты способны взаимодействовать с бактериофагами, так как содержат остатки пептидогликанового слоя. После удаления ингибиторов, вызвавших нарушение синтеза клеточной стенки, измененные бактерии реверсируют в исходное состояние, то есть восстанавливают полноценную клеточную стенку и первоначальную форму клеток.
Бактерии, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L- формами. L-формы бактерий представляют собой осмотически чувствительные шаровидные или колбовидные клетки различной величины.
Различают стабильные и нестабильные L-формы. Стабильные L-формы не способны к реверсии в исходные бактериальные клетки. Нестабильные L-формы возвращаются в исходную бактериальную форму после удаления фактора, приведшего к изменению бактерий. L-формы могут образовывать многие возбудители инфекционных болезней, в том числе в организме человека или животных. Образование L-форм бактерий называется L-трансформацией.
У некоторых микроорганизмов клеточная стенка имеет особенности строения. В частности, у хламидий клеточная стенка состоит из внутренней цитоплазматической мембраны и внешней мембраны. Каждая мембрана является двойной. В отличие от других грамотрицательных бактерий, клеточная стенка хламидий не имеет пептидогликанового слоя. В состав клеточной стенки хламидий входят пептиды и гликолипиды (аналоги липополисахаридов клеточной стенки грамотрицательных бактерий). Основными белками клеточной стенки хламидий являются белки внешней мембраны ОМР 2 и МОМР (рисунок 45).
Цитоплазматическая мембрана бактерий состоит из двойного слоя фосфолипидов (бимолекулярный липидный слой) и мембранных белков (рисунок 46).
Поры между белками
Слой белков
Цитоплазматическая мембрана образует барьер, препятствующий движению веществ внутрь клетки и наружу. Мембранные белки подразделяются на поверхностные и погруженные (интегральные, тоннельные). Среди мембранных белков особую роль выполняют пермеазы, участвующие в транспорте веществ внутрь клетки (транспортная функция цитоплазматической мембраны).
Цитоплазматическая мембрана окружает цитоплазму бактерий и участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки. Она играет значительную роль в процессах роста и деления клеток, в движении бактерий, в секреции веществ, в спорообразовании, то есть в процессах с высокой затратой энергии.
У многих бактерий в зоне формирования поперечных перегородок при делении клеток цитоплазматическая мембрана образует впячивания (инвагинаты, дивертикулы) в виде сложных мембранных структур. Эти структуры называются мезосомами. Они имеют форму цистерн, пузырьков, канальцев.
Выделяют три типа мезосом: ламеллярные (пластинчатые), везикулярные (имеющие форму пузырьков) и тубулярные (трубчатые). Эти мезосомы различаются по строению. Часто наблюдаются мезосомы смешанного типа. По своему расположению в клетке выделяют септальные мезосомы (располагаются в зоне клеточного деления и формирования поперечной перегородки или септы), латеральные (инвагинации периферических участков цитоплазматической мембраны) и мезосомы, к которым прикреплен нуклеоид. Мезосомы выполняют функцию генерации энергии, участвуют в процессах роста и деления клеток, в синтезе углеводов, липидов и других компонентов клетки.
Цитоплазма является основной центральной частью клетки. Она отграничена цитоплазматической мембраной Цитоплазма представляет собой коллоидную систему (цитозоль), состоящую из растворимых белков, рибонуклеиновых кислот, включений запасных органических веществ и многочисленных мелких гранул - ри- босом. Включения придают цитоплазме мелкозернистый вид.
Рибосомы - это немембранные органоиды бактериальной клетки. Они служат для биосинтеза белка из аминокислот, находящихся в цитоплазме, на основе информации, предоставляемой матричной РНК (мРНК).
Рибосомы представляют собой мелкие структуры сферической или эллипсоидной формы. Они имеют размер около 10-20 нм. Бактериальные рибосомы состоят из двух субъединиц - малой (30S) и большой (50S). Они представляют собой нуклеопротеид, в составе которого 60% составляет РНК и 40% - белок. Рибосомные РНК (рРНК) являются консервативными элементами бактерий.
В цитоплазме бактерий находятся различные включения в виде гранул полисахаридов, жировых соединений и полифосфатов. Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей бактерий. Гранулы полисахаридов у одних бактерий содержат крахмал (нейссерии), у других – гликоген (бациллы, клостридии), у третьих - гранулезу (клостридии).
Жировые включения состоят из поли-β-оксимасляной кислоты, нейтральных жиров и жировосковых веществ (у микобактерий, грибов). Гранулы полифосфатов (волютин) являются запасным резервуаром фосфатов, необходимых при синтезе АТФ и ДНК (у коринебактерий, микобактерий, актиномицетов). Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки.
Нуклеоид является генетическим аппаратом бактерий (эквивалент ядра эукариотической клетки). Он расположен в центральной зоне бактериальной клетки и представляет собой двунитевую молекулу ДНК, замкнутую в кольцо и плотно уложенную наподобие клубка. ДНК в развернутом состоянии имеет длину более 1 мм. Нуклеоид бактерий, в отличие от ядра эукариотических клеток, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК методами: по Фельгену или по Романовскому-Гимзе. При электронной микроскопии ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой.
Кроме нуклеоида в цитоплазме бактериальной клетки могут находиться внехромосомные ковалентно замкнутые кольцевые молекулы ДНК или плазмиды
Некоторые плазмиды могут быть интегрированы с бактериальной хромосомой. Плазмиды придают бактериальной клетке определенные селективные преимущества: устойчивость к антибиотикам (R-плазмиды), продуцирование бактериоцинов (Col-плазмиды), синтез токсинов (Tox-плазмиды) и др. Плазмиды, свойства которых не установлены, называются криптическими.
Капсула является надоболочечной структурой клетки, имеющей слизистую консистенцию. Толщина капсулы составляет более 0,2 мкм. Она прочно связана с клеточной стенкой бактерий и имеет четко очерченные внешние границы. Различают макрокапсулу (собственно капсулу) и микрокапсулу. Макрокапсула выявляется в световом микроскопе в виде неокрашенной зоны, окружающей клетку. Капсула хорошо различима в мазках-отпечатках из патологического материала и в мазках, приготовленных из чистых культур бактерий.
В чистых культурах бактерий капсула выявляется путем окраски препарата по методу Бурри-Гинса. При этом используют тушь и раствор фуксина. Тушь создает темный фон вокруг капсулы, а бактерии окрашиваются фуксином в красный цвет
Капсула состоит из полисахаридов (экзополисахаридов), гликопротеинов, полипептидов. Например, у возбудителя сибирской язвы капсула состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, включает большое количество воды. Капсула препятствует фагоцитозу бактерий в организме, способствует адгезии бактерий к субстратам, предохраняет бактерии от высыхания. Капсула обладает антигенностью: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы). Капсульные бактерии на плотных питательных средах формируют гладкие блестящие колонии слизистой консистенции. Утрата капсулы снижает патогенность бактерий.
Многие бактерии образуют микрокапсулу - слизистое образование на поверхности клетки толщиной менее 0,2 мкм. Микрокапсула выявляется только с помощью электронной микроскопии.
У некоторых бактерий на поверхности клеток обнаруживается слизистый слой. Он не имеет четких внешних границ и прочной связи с клеткой, поэтому легко от нее отделяется. Слизистый слой не виден при световой микроскопии. Он выявляется серологическими методами или при электронной микроскопии.
Жгутики являются надоболочечными структурами бактериальной клетки. Они выполняют функцию органа движения бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны. Длина жгутиков значительно превышает размеры клетки. Жгутики выявляются при световой микроскопии только после специального окрашивания: серебрением по Морозову, окраской по Грею. Наиболее четко жгутики выявляются при электронной микроскопии при напылении тяжелыми металлами (рисунок 55).
Жгутик состоит из 3 частей:
- базальное тельце;
- крюк (колено);
- спиралевидная нить (филамент, собственно жгутик).
Базальное тельце включает в себя стержень с системой дисков и белки мотора. У грамположительных бактерий имеется одна пара дисков, а у грамотрицательных бактерий - две пары дисков. Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. Базальное тельце является своего рода электромотором, вращающим жгутик (рисунок 56).
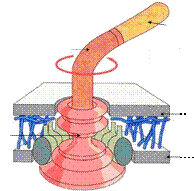
Филамент
Крюк
Базальное тельце
Наружная мембрана
ЦПМ
В качестве источника энергии при движении жгутиков используется разность потенциалов на цитоплазматической мембране. Скорость вращения жгути- ка может достигать 100 об/с. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер. Жгутики вращаются по часовой или против часовой стрелки. В зависимости от этого клетка движется либо вперед, либо назад.
Жгутики состоят из особого белка флагеллина (flagellum - жгутик). Этот белок обладает высокой антигенной активностью (Н-антиген бактерий). Субъединицы флагеллина закручены в виде спирали. Количество и расположение жгутиков у разных видов бактерий варьирует. Бактериальная клетка может содержать до 1000 жгутиков. В зависимости от количества и локализации жгутиков выделяют следующие группы бактерий (рисунок 57):
- монотрихи (греч. monos - один, trichos - волос) - бактерии, имеющие один жгутик, например, холерный вибрион (рисунок 58);
- лофотрихи (греч. lophos - пучок, trichos - волос) - бактерии, имеющие пучок жгутиков на одном из концов клетки, например, кампилобактерии (рисунок 59);
- амфитрихи (греч. amphi - с обеих сторон, trichos - волос) - бактерии, имеющие по одному жгутику или пучку жгутиков на противоположных концах клетки, например, спириллы (рисунок 60);
- перитрихи (греч. peri - около, trichos - волос) - бактерии, имеющие большое количество жгутиков, покрывающих всю поверхность клетки, например, кишечная палочка (рисунок 61).
Рисунок 57 - Расположение жгутиков у бактерий: А- монотрихиальное; В – лофотрихиальное; С – амфитрихиальное; D – перитрихиальное.
Пили (фимбрии, ворсинки) - это нитевидные выросты на поверхности клетки. Пили располагаются либо перитрихиально, либо локализуются на одном из концов клетки. Пили берут начало от цитоплазматической мембраны и состоят из белка пилина. Белковые субъединицы закручены вокруг полой сердцевины. Пили встречаются как у подвижных, так и неподвижных бактерий. Они обладают антигенностью. Различают общие пили или пили первого типа (пили-адгезины, отвечают за адгезию бактерий к различным субстратам) и половые пили, пили второго типа или конъюгативные пили (F-пили). Общие пили детерминируются хромосомными генами, а конъюгативные пили - внехромосомным фактором фертильности (F-плазмидой).
Жгутики
Капсула
Включения Рибосомы
Нуклеоид
Плазмида
Мезосома
Клеточная стенка
Цитоплазматическая мембрана
Бактериальная оболочка состоит из клеточной стенки и располагающейся под ней цитоплазматической мембраны. Клеточная стенка - это ригидная структу- ра, которая придает бактериальной клетке определенную форму. Она защищает внутреннее содержимое клетки от вредных воздействий внешней среды, участвует в процессах деления клетки и транспорта метаболитов. На поверхности клеточной стенки располагаются рецепторы для бактериофагов, бактериоцинов, антибиотиков и других химических веществ. По строению клеточной стенки различают фирмикутные бактерии (грамположительные, толстостенные), грациликутные бактерии (грамотрицательные, тонкостенные) и бактерии, не имеющие клеточной стенки (микоплазмы). Подразделение бактерий на грамположительные (грампозитивные) и грамотрицательные (грамнегативные) основано на разном восприятии красителей при окраске по методу, предложенному датским бактериологом Г. К. Грамом.
Клеточная стенка грамположительных бактерий представляет собой гомогенный слой. Она состоит из многослойного пептидогликана (муреина), пронизанного молекулами тейхоевой и липотейхоевой кислот.
Пептидогликан клеточной стенки образован параллельно расположенными молекулами гликана, состоящего из остатков N-ацетилглюкозамина и N- ацетилмурамовой кислоты, соединенных вдоль гликозидной связью.
Тейхоевые кислоты представляют собой цепи из остатков глицерола и рибитола, соединенных фосфатными мостиками. Пептидогликан и тейхоевые кислоты в конечном итоге формируют так называемый муреиновый мешок,покрывающий клетку снаружи. Тейхоевые кислоты позволяют муреиновому мешку растягиваться и сжиматься, действуя наподобие пружин. Тейхоевые кислоты выполняют антигенную и адгезивную функции грамположительных бактерий. Пептидогликан грамположительных бактерий плотно прилегает к цитоплазматичекой мембране. Клеточная стенка грамположительных бактерий содержит также небольшое количество полисахаридов, белков и липидов. В клеточной стенке имеются поры диаметром 1-6 нм, через которые внутрь клетки проникают различные вещества.
При окраске по Граму толстый слой пептидогликана грамположительных бактерий удерживает генциановый фиолетовый в комплексе с йодом. Последующая обработка препарата спиртом вызывает суживание пор в пептидогликане и тем самым усиливает задержку фиолетового красителя в клеточной стенке. Заключительная окраска препарата фуксином не изменяет первоначальной окраски клеток. Грамположительные бактерии окрашиваются в сине-фиолетовый цвет.
Клеточная стенка грамотрицательных бактерий выделяют внешнюю (наружную) мембрану (НМ) и тонкий пептидогликановый слой или муреиновый мешок (рисунок 37).
Внешняя мембрана грамотрицательных бактерий представляет собой фосфолипидный бислой, содержащий белки и липополисахарид. Липополисахарид (ЛПС) состоит из трех составных частей:
- липид А - специфический гликолипид, встроенной в фосфолипидный бислой, закрепляющий молекулу ЛПС во внешней мембране и придающий липополисахариду токсические свойства;
- ядро - центральная (стержневая, коровая) область (лат. core - ядро) полисахаридной природы;
- боковая О-цепь, образованная повторяющимися олигосахаридами (О- антиген).
Боковая О-цепь
Ядро
Липид А
С помощью мембранного липопротеина внешняя мембрана связана с подлежащим слоем пептидогликана. Пептидогликан грамотрицательных бактерий является однослойным (толщина – 2-3 нм) и не содержит тейхоевых кислот. Под слоем пептидогликана располагается цитоплазматическая мембрана. Между внешней мембраной, слоем пептидогликана и цитоплазматической мембраной имеется полость, называемая периплазматическим пространством (периплазмой). Это пространство заполнено гелем, содержащим транспортные белки и ферменты (протеазы, липазы, фосфатазы, нуклеазы, бета- лактамазы). В периплазматическом геле располагается муреиновый (пептидогликановый) мешок.
Белки наружной мембраны включают порины, трансмембранные белки и белки, участвующие в формировании поверхностных структур (пилей, жгутиков). Порины образуют каналы для проникновения воды и мелких молекул. Трансмембранные белки обеспечивают связь с муреиновым мешком.
При окраске по Граму тонкий слой пептидогликана грамотрицательных бактерий под воздействием этилового спирта утрачивает комплекс генцианового фиолетового и йода. При последующей обработке фуксином или сафранином клетки приобретают красный цвет.
Рисунок 40 - Клеточные стенки грамположительных и грамотрицательных
бактерий.
К грамположительным бактериям относятся все кокки (за исключением гонококков и менингококков), спорообразующие палочки (бациллы и клостридии), микобактерии, коринебактерии, листерии. К грамотрицательным бактериям относятся гонококки и менингококки, не образующие спор палочки, извитые бактерии.
Клеточная стенка бактерий выполняет следующие функции:
- предохраняет клетку от вредных воздействий окружающей среды;
- обеспечивает постоянство формы клетки;
- сообщает бактериальной клетке антигенные свойства;
- регулирует рост и деление клетки;
- участвует в поступлении внутрь клетки некоторых молекул;
- обеспечивает тинкториальные свойства бактерий (отношение к красителям).
Под воздействием некоторых веществ (лизоцим, пенициллин, гуморальные факторы организма) нарушается синтез компонентов клеточной стенки бактерий. В таких случаях бактерии полностью или частично лишаются клеточной стенки, образуя шаровидные формы. Такие формы имеют размеры, превышающие исходные клетки в несколько раз. Бактерии, полностью лишенные клеточной стенки, называются протопластами, а бактерии, частично сохранившие клеточную стенку, называются сферопластами.
Образование протопластов характерно для грамположительных бактерий. Протопластообразование сопровождается утратой толстой пептидогликановой клеточной стенки. Протопласты содержат только цитоплазматическую мембрану (рисунок 42). Для их поддержания требуется изотоническая среда. Они устойчивы к антибиотикам и бактериофагам.
Клеточная стенка
Цитоплазматическая мембрана
Сферопласты образуются грамотрицательными бактериями. Образование сферопластов сопровождается утратой внешней мембраны клеточной стенки. Но сферопласты наряду с цитоплазматической мембраной содержат тонкий слой пептидогликана (рисунок 43).
Пептидогликан
Клеточная стенка
Цитоплазматическая мембрана
Рисунок 43 – Образование сферопластов у бактерий.
Для поддержания сферопластов также требуется среда с повышенным осмотическим давлением. Сферопласты способны взаимодействовать с бактериофагами, так как содержат остатки пептидогликанового слоя. После удаления ингибиторов, вызвавших нарушение синтеза клеточной стенки, измененные бактерии реверсируют в исходное состояние, то есть восстанавливают полноценную клеточную стенку и первоначальную форму клеток.
Бактерии, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L- формами. L-формы бактерий представляют собой осмотически чувствительные шаровидные или колбовидные клетки различной величины.
Различают стабильные и нестабильные L-формы. Стабильные L-формы не способны к реверсии в исходные бактериальные клетки. Нестабильные L-формы возвращаются в исходную бактериальную форму после удаления фактора, приведшего к изменению бактерий. L-формы могут образовывать многие возбудители инфекционных болезней, в том числе в организме человека или животных. Образование L-форм бактерий называется L-трансформацией.
У некоторых микроорганизмов клеточная стенка имеет особенности строения. В частности, у хламидий клеточная стенка состоит из внутренней цитоплазматической мембраны и внешней мембраны. Каждая мембрана является двойной. В отличие от других грамотрицательных бактерий, клеточная стенка хламидий не имеет пептидогликанового слоя. В состав клеточной стенки хламидий входят пептиды и гликолипиды (аналоги липополисахаридов клеточной стенки грамотрицательных бактерий). Основными белками клеточной стенки хламидий являются белки внешней мембраны ОМР 2 и МОМР (рисунок 45).
Цитоплазматическая мембрана бактерий состоит из двойного слоя фосфолипидов (бимолекулярный липидный слой) и мембранных белков (рисунок 46).