Лекция №1
Структура белков, углеводов и липидов.
План.
1. Структура белков.
2. Стрктура углеводов.
3. Структура липидов.
Структура белков.
Белки - органические соединения, состоящие из аминокислот, связанных между собой пептидной связью. Аминокислоты - кабоновые кислоты, содержащие аминную и карбоксильную групп, находящиеся у одного и того же углеродного атома.В состав белков входят 22 аминокислоты, которые по свойствам боковой цепи делят на 4 группы: гидрофобные,
гидрофильные неионные, гидрофильные с отрицательным зарядом и гидрофильные с положительным зарядом.
I. гидрофобные аминокислоты.



Глицин. Аланин Валин Лейцин

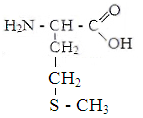


Изолейцин Метионин Цистин Фенилаланин


Триптофан Пролин
II. гидрофильные неионные аминокислоты.



Серин Треонин Аспарагин Глутамин

Оксипролин
III. гидрофильные аминокислоты.
.


Аспарагиновая Глутаминовая Цистеин Тирозин
кислота кислота
IV. гидрофильные аминокислоты с положительно заряженным радикалом.



Лизин Аргинин Гистидин
Биологическая роль углеводов.
Углеводы являются основным энергетическим материалом в организме. При окислении 1 г глюкозы высвобождается 4,1 ккал. Углеводы необходимы для окисления липидов. Гликоген и крахмал являются запасным энергетическим материалом.
Рибоза входит в состав нуклеотидов РНК, АТФ, НАД, ФАД, витамина В12 и других веществ, а дезоксирибоза - в состав ДНК.
Углеводы являются источником пластического материала для биосинтеза триглицеридов, холестерина, заменимых аминокислот и других веществ.
Гетерополисахариды являются небелковой частью гликопротеинов, участвующих в многих процессах: формировании соединительной ткани, развитии иммунной реакции, свертывании крови и др.
Лекция № 2
ФЕРМЕНТЫ
План
1.Структура ферментов
2. Свойства ферментов.
Структура ферментов
Ферменты (энзимы) – белки, являющиеся биологическими катализаторами. По строению могут быть простыми и сложными белками. Последние состоят из белковой части (апофермента) и небелковой части (кофермента). По отдельности каталитической активностью не обладают, активны лишь вместе. Существуют мультиферментные комплексы, состоящие из нескольких ферментов
Некоторые ферменты имеют четвертичную структуру – состоят из двух и более радикалов более мелких белков (субъединиц). Некоторые ферменты могут катализировать одну реакцию, но отличаться по строению субъединиц.
Их называют изоферментами. Например, креатинкиназа-1 мозга состоит из субъединиц ВВ, креатинкиназа-2 миокарда - из субъединиц ВМ, а креатинкиназа-3 скелетных мышц – из субъединиц ММ.
В молекуле фермента различают активый и аллостерический центры. К первому из них присоединяется вещество, на которое действует фермент (субстрат), происходит его превращение, образуется продукт реакции. Этот центр занимает лишь часть молекулы фермента. В его составе выделяют контактный участок, к которому присоединяется субстрат и каталитический центр, где происходит ферментативная реакция. В составе каталитического центра могут быть радикалы аминокислот, содержащие группы –СООН, -ОН, -NH2 и др., а у ферментов-сложных белков – и кофермент.
Аллостерический центр (allos – другой) – участок фермента, к которому присоединяются вещества, отличные от субстрата, способные изменять форму активного центра, ускоряя таким образом ферментативную реакцию (активаторы) или замедляя ее (ингибиторы). Такими веществами могут быть продукты собственной реакции, других реакций, гормоны и др.
Строение коферментов.
Различают коферменты витаминные т невитаминные.
Витаминные коферменты.. К ним относят никотинамидные, флавиновые, тиаминовые, пиридоксалевые, пантотеновые, кобаламиновые и другие коферменты.
Никотинамидные коферменты – производные витамина РР.
К ним относят никотинамидадениндинуклеотид (НАД) и никотинамидадениндинуклеотид фосфат (НАДФ). НАД из соединенных между собой остатков аденозинмонофосфата (АМФ) и никотинамидрибозилмонофосфата (НМФ). Он входит в состав ферментов дегидрогеназ, отщепляющих ионы водорода от гидроксильных, альдегидных или аминогрупп. Ионы водорода присоединяются к атому углерода никотинамида, находящемуся в четвертом положении. НАД способен переносить ионы водорода в дыхательную цепь митохондрий, где они окисляются с образованием АТФ.

НАДФ отличается по строению тем, что к второму углеродному атому радикала рибозы АМФ присоединяется остаток фосфорной кислоты. Является коферментом дегидрогеназ пентозного цикла, откуда ионы водорода поставляются в реакции биосинтеза различных веществ.
Флавиновые коферменты – производные витамина В2.
К ним относя флавинмононуклеотидд (ФМН) и флавинадениединуклеотид (ФАД). Первый из них входит в состав дыхательной цепи митохондрий. Он принимает ионы водорода и электроны от НАД-Н2 и передает их убихинону.

ФАД состоит из остатков АМФ и ФМН. Является коферментом дегидрогеназ, отщепляющих ионы водорода от янтарной кислоты и активированных жирных кислот. Ионы водорода присоединяются к атомам азота изоалаксозинового цикла и переносятся в дыхательную цепь митохондрий.
Тиаминовые коферменты – производные витамина В1. К ним относя тиаминдифосфат (ТДФ). Входит в состав транскетолазы пентозного цикла, а также ферментов отщепляющих углекислый газ от пировиноградной и альфа-кетоглутаровой кислот.

Пиридоксальфосфат (ПФ) – производное витамина В6. Входит в состав ферментов обмена аминокислот – декарбоксилаз, трансамназ.

Пантотеновые коферменты – производные витамина В3. К ним относят коэнзим А (КоА), входящий в состав ферментов, переносящих остатки уксусной, янтарной или жирных кислот.
СН3 ОН
!!
АДФ-О - СН2 - С----СН – СО – NH - СН2 -СН2 - COOH
!
СН3
Коэнзим А
Фолатные коферменты - производные витамина Вс (фолиевой кислоты).

Участвуют в переносе метильных, альдегидных и других одноуглеродных фрагментов
Невитаминными коферментами могут быть гем (у каталазы, цитохромов и др. ферментов), ионы металлов (у глутатионпероксидазы, супероксиддисмутазы, амилазы и др.), глутатион (у глицеральдегид-3-фосфатдегидрогеназы), мононуклеотиды (АМФ, ГМФ, ЦМФ) и др.

Свойства ферментов
1. Высокая каталитическая активность – способность повышать скорость реакций в миллионы-миллиарды раз. Неорганические катализаторы увеличивают ее лишь в тысячи раз.
2. На активность фермента влияет концентрация субстрата. При увеличении последней активность фермента возрастает, но до определенного предела. Концентрация субстрата, при которой активность фермента равна 0,5 от максимальной, называется константой Михаэлиса.
3. На активность фермента влияет концентрация продукта реакции. При постепенном накоплении последнего в среде она снижается. Поэтому активность фермента определяют на начальном этапе реакции, когда субстрат еще не накопился.
4. Активность ферментов зависит от рН среды. В зависимости от рН изменяется диссоциация ионогенных групп в активном центре фермента и в молекуле субстрата, что влияет на скорость реакции. Большинство ферментов наиболее активны в слабощелочной среде. Исключение – пепсин желудочного сока и катепсины лизосом, активные в кислой среде.
5. На активность фермента влияет температура среды. Обычно при повышении ее на 10оС скорость реакции увеличивается в 2 раза (правило Ван Гоффа). Наиболее активны ферменты млекопитающих при 40-50 о. При более низкой температуре активность тормозится из-за снижения диссоциации ионогенных групп, а при более высокой – из-за денатурации белковой молекулы.
6. Ферменты обладают специфичностью, т.е. способность воздействовать лишь на определенный субстрат. Различают различные виды специфичности.
а. Абсолютная специфичность, когда фермент действует лишь на один субстрат. Например, фумаратгидратаза действует только на фумаровую кислоту.
б. Групповая специфичность, когда фермент действует на несколько сходных по структуре субстратов. Например, липаза расщепляет ацилглицеролы, состоящие из различных жирных кислот.
г. Стереоспецифичность, когда фермент действует только на определенный пространственный изомер. Например, фумаратгидратаза не действе на стереоизомер фумаровой кислоты – малеиновую кислоту.
7. На активность ферментов влияют вещества, повышающие скорость реакции (активаторы) и снижающие ее (ингибиторы). Некоторые ферменты вырабатываются неактивными и при помощи активаторов переходят в активные. Например, энтерокиназа отщепляет от трипсиногена часть молекулы и он превращается в активный трипсин. Активаторами могут быть ионы металлов, субстраты, продукты других реакций, гормоны и другие вещества.
Ингибиторы по характеру торможения активности фермента могут быть обратимыми и необратимыми. В первом случае комплекс фермент-ингибитор может расщепляться и активность фермента восстанавливается. При необратимом ингибировании образуется прочная связь между ферментом и ингибитором.
По механизму действия ингибиторы делят на конкурентные, неконкурентные, субстратные и аллостерические.
а. При конкурентном ингибировании структура ингибитора близка к структуре субстрата и он конкурирует за активный центр. Например, малоновая кислота по структуре близка к янтарной кислоте и поэтому способна связывать активный центр сукцинатдегидрогеназы. Ингибирование можно ослабить увеличением концентрации субстрата. На конкурентном ингибировании основано действие сульфаниламидов. Они по строению сходны с п-аминобензойной кислотой, необходимой микроорганизмам. Сульфаниламиды, занимая место последней, тормозят рост микроорганизмов.
б. Неконкурентное ингибирование развивается под действием вещества, структура которого не сходная со структурой субстрата. Оно связывается не с активным центром, а с другими участками молекулы фермента. Последний после этого может образовывать комплекс с субстратом, но внутри этого комплекса реакция не протекает. Например, ионы тяжелых металлов связывают HS-группы ферментов, ингибируя их. Цианиды связываются с ионом железа цитохромоксидазы, прекращая дыхание митохондрий. Неконкурентное ингибирование, в отличие от конкурентного, не снимается избытком субстрата.
в. Субстратное ингибирование развивается иногда при увеличении в среде концентрации субстрата. Его снимают, уменьшая концентрацию последнего.
г. Аллостерическое ингибирование развивается тогда, когда вещество, сзязываясь с аллостерическим центром, изменяет конфигурацию активного центра, блокируя его. Такими ингибиторами могут быть собственный продукт реакции, продукты других реакций, гормоны и другие вещества.
Лекция №3.
ФЕРМЕНТЫ (продолжение)
План.
1. Механизм действия Ф.
2.Номенклатура и классификация ферментов.
3. Ферменты в сельском хозяйстве.
Механизм действия ферментов
Ферменты во много раз увеличивают скорость химических реакций, что с термодинамических позиций связывают со способностью их снижать энергию активации реакции, т. е. то дополнительное количество ее, которое нужно дать молекулам, чтоб они из неактивного состояния перешли в активное. Это снижение происходит через несколько стадий.
Согласно теории Михаэлиса-Ментена ферментативная реакция протекает в три стадии: 1) фермент (Фер) + субстрат (Суб) ------ фермент-субстратный комплекс (Фер-Суб);
2) преобразование субстрата в несколько стадий молекулы внутри фермент-субстратного комплекса с образованием внутри него промежуточных продуктов и в конечном итоге - комплекса фермент-продукт реакции (Фер- Прод); 3) расщепление комплекса фермент-продукт реакции с диффузией фермента и продукта реакции в среду: Фер-Прод ------ Фер + Прод.
На первой стадии реакции субстрат маневрирует, сближаясь с активным центру фермента, концентрируясь в нем. При этом происходит их взаимная пространственная подгонка так, чтоб активный центр расположился напротив «атакуемой» связи в молекуле субстрата. Образуется фермент-субстратный комплекс. Процесс протекает мгновенно, энергия активации снижается незначительно.
Вторая стадия реакции наиболее продолжительная, за счет нее в основном снижается энергия активации. В активном центре фермента и в молекуле субстрата содержатся кислотные и основные радикалы. Поэтому в ходе реакции может изменяться рН среды, что влияет на ее скорость. Во время нее происходит натяжение, деформирование химических связей в молекуле субстрата, их разрыв с возникновением новых связей. Это приводит к ускорению реакции, образования во время нее вначале одного или нескольких промежуточных продуктов, затем - продукта реакции.
Третья стадия реакции протекает также мгновенно и энергия активации изменяется мало.
Лекция №4
ОБМЕН ВЕЩЕСТВ
План
- Понятие об обмене веществ.
- Биологическое окисление.
- Обмен энергии. АТФ. Митохондрии.
- Цикл трикарбоновых кислот.
Обмен веществ – совокупность химических превращений, которым подвергаются вещества с момента поступления в организм до выделения их в виде конечных продуктов. Включает два взаимосвязанные между собой процессы: ассимиляцию и диссимиляцию. Ассимиляция – образование характерных для организма веществ, сопряженное с потреблением АТФ. Диссимиляция – процесс распада веществ в организме.
Этапы обмена веществ
- Пищеварение – гидролитическое расщепление ферментами веществ,
поступивших с пищей, до простых веществ- мономеров: белки расщепляются до аминокислот, жиры – до глицерина и жирных кислот, олиго- и полисахахариды – до моносахаридов.
- Всасывание мономеров в кровь в стенке кишечника.
- Межуточный (тканевой) обмен – химические превращения веществ в
тканях или метаболизм. Вещества, участвующие в этих превращениях – метаболиты. Одновременно в организме протекают процессы образования (анаболизм) и распада веществ (катаболизм).
4. Образование конечных продуктов -процесс, позволяющий высвободить около 80% энергии. Конечными продуктами метаболизма белков, углеводов и липидов являются углекислый газ и вода. При окислении белков образуется также аммиак, обезвреживающийся в мочевину.
Биологическое окисление – совокупность процессов окисления (отщепления и присоединения электронов), протекающих в клетках. Согласно теории В.И. Палладина оно заключается в отщеплении от окисляемого субстрата ионов водорода и электронов и переносе их на активированный кислород с образованием воды:
S-H2 2Н+ + 2е- ½ О2 → Н2О + энергия (210 кДж).
Реакция 2Н+ + ½ О2 → Н2О + энергия является основным источником энергии в организме. Вне организма она протекает быстро, с выделением большого количества энергии, с взрывом, а в организме – ступенчато. Энергия при этом выделяется порциями, что предотвращает перегревание клетки. Часть ее выделяется в виде тепла, часть запасается в макроэргических связях аденозинтрифосфата (АТФ):
Аденозиндифосфат (АДФ) + Н3РО4 + энергия → АТФ.
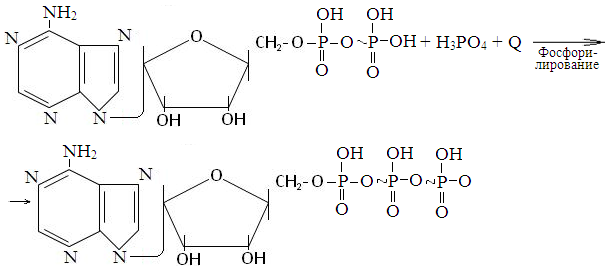
Синтез АТФ из АДФ и Н3РО4 называется фосфорилированием. Различают две разнолвидности его: субстратное и окислительное фосфорилирование. Первое из них (субстратное фосфорилирование) протекает за счет энергии, освобождающейся при расщеплении макроэргических (богатых энергией) химических связей некоторых веществ- субстратов: 1,3-дифосфоглицериновой, фосфоенолпировиноградной кислот (в реакциях окисления глюкозы) и активной формы янтарной кислоты (в реакциях окисления активной формы уксусной кислоты в цикле Кребса); 2) за счет энергии освобождающейся при окислении кислородом ионов водорода ферментами дыхательной цепи, расположенной на внутренней мембране митохондрий (окислительное фосфорилирование).
Строение дыхательной цепи митохондрий
На внутренней мембране этих органоидов локализовано 8 оксидоредуктаз, коферменты которых располагаются последовательно в следующем порядке:
Ц и т о х р о м ы
S-H2 2е- НАД 2е- ФМН 2е- убихинон 2е- Ц. b 2е- Ц. c1 2е- Ц. c 2е- Ц. a 2е- Ц. a3 2е-
2Н+ 2Н+ 2 Н + 2Н+
Н 2О ½ О2
-------------------- ----------------- -----------------
АТФ АТФ АТФ
Первый кофермент – НАД. Он принимает ионы водорода и электроны от субстрата S-H2, находящегося за пределами дыхательной цепи. Акцептором их является атом углерода никотинамида, находящийся в 4-м положении. Образовавшийся НАД-Н2 передает ионы водорода и электроны ФМН, где они присоединяются к атомам азота изоалаксозинового цикла. От ФМН-Н2 ионы водорода и электроны переходят на убихинон. Первые из них временно остаются на убихиноне и в дальнейшем, соединяясь с атомом активированного кислорода, образуют молекулу воды.
От убихинона електроны передаются на цитохром b, присоединяясь к атому железа гема, который меняет свою валентность с 3 на 2.. От цитохром b они перемещаются последовательно на цитохромы с1, с, а и а3. При каждом таком переходе электрона от одного фермента дыхательной цепи митохондрий к другом происходит постепенная потеря энергии и на цитохром а3 (цитохромоксидазу) переходит лишь остаток ее. Последний передается на молекулу кислорода, поступившую в клетку через кровь из легких, активируя ее. Активированный кислород после этого соединяется с ионами водорода, отщепившимися от убихинона и образуется вода, конечный продукт биологического окисления.
Генерации АТФ ферментами дыхательной цепи митохондрий
Каждые два фермента дыхательной цепи митохондрий образуют редокс-пару. Между ними имеются определенный окислительно-восстановительный потенциал (ОВП), э.д.с., выражающаяся в вольтах. В процессе переноса электронов происходит постепенное увеличение ОВП. У НАД он равен -0,32 в, у кислорода - +0,81 в. Генерация АТФ в дыхательной цепи митохондрий происходит на тех участках ее, где различие в ОВП между компонентами редокс-пары составляет не менее 0,2 в. Таких участков 3: 1) между НАД и ФМН; 2) между цитохромами b и c1; 3) между цитохромами а и а3.Поэтому при окислении ферментами дыхательной цепи митохондрий одной молекулы НАД-Н2 генерируется 3 молекулы АТФ. Если же окисляется ФАД-Н2, то ионы водорода и электроны передаются от него на ФМН, минуя первый пункт генерации АТФ, между НАД и ФМН. Поэтому произойдет генерация только двух молекул АТФ.
Согласно теории П. Митчелла, энергия, отдаваемая электронами при прохождении через эти участки дыхательной цепи, используется для перемещения положительно заряженных ионов водорода из внутреннего пространства митохондрий (матрикса) в пространство между внутренней и наружной мембранами их, т.е. на наружную поверхность внутренней мембраны этих органоидов (ВММ). Вследствие этого на последней накапливается избыток ионов водорода. При этом на внутренней поверхности ВММ содержится избыток отрицательно заряженных электронов. Поэтому создается разница потенциалов между наружной и внутренней поверхностями ВММ (мембранный потенциал).
Из-за наличия этого потенциала ионы водорода стремятся вернуться обратно в матрикс. Для этого они использую специальный канал, расположенный внутри фермента протонная АТФ-аза, расположенная на ВММ. Во время прохождения через этот канал ионов водорода выделяется энергия, которая используется для генерации из АДФ и фосфорной кислоты АТФ (окислительного фосфорилирования). Следовательно протонная АТФ-аза функционирует как АТФ- синтетаза.
Креатинкиназная реакция – «челночный» механизм, осуществляющий перенос энергии АТФ от места выработки его (митохондрий, расположенных на поверхности мышечных волокон) до места потребления (саркомеров мышечных волокон). Прямо поступать АТФ в саркомеры не может, т.к. расщепляется по пути ферментами.
Микросомальное окисление, протекающее с участие специальной цепи переноса электронов (ЦПЭ), расположенной на мембранах эндоплазматической цепи печени, надпочечников и других органов, также сопряжено с потреблением кислорода. При этом последний не только превращается в воду, но и используется для гидроксилирования (присоединения НО-группы) ряда веществ: стероидных гормонов, ядов лекарственных и других веществ, происходящее на первой стадии их инактивации. Микросомальная ЦПЭ состоит из четырех ферментов, коферментами которых являются НАДФ, ФАД, ион железа и цитохром Р450:
О R-H
2 Н+
НАДФ-Н2 2Н+ флавопртеин --- Fe-cодержащий белок -- цит. Р450 --О R-OH
2е- 2е- 2е- 2е-
Источники ионов водорода в клетках.
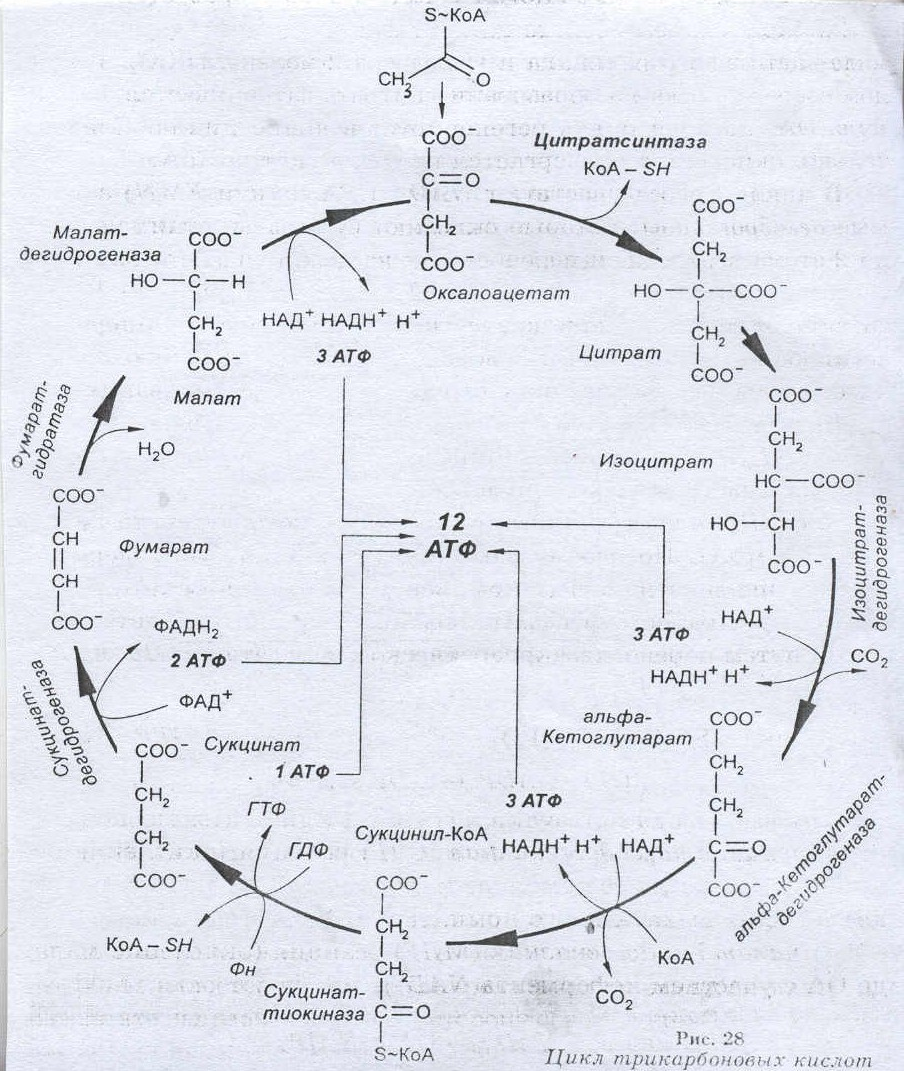
Большое количество ионов водорода генерируются в реакциях отщепления их от 3-фосфоглицеринового альдегида, молочной, пировиноградной кислот (окисление глюкозы), активированных жирных кислот (окисление липидов), глутаминовой кислоты (обмен белков). Основная масса их высвобождается при окислении в цикле трикарбоновых кислот, описанном Г. Кребсом, активированной уксусной кислоты (ацетил-КоА). Источником последнего является главным образом окисление пировиногроадной, жирных кислот и некоторых аминокислот.
Лекция №5
Цикл Кребса
План.
1. Реакции цикла трикарбоновых кислот.
2. Биологическая роль цикла Кребса.
Цикл Кребса – 10 последовательно протекающих реакций, начинающихся с вовлечения в процесс шавелевоуксусной кислоты и заканчивающихся высвобождением ее.
Лекция №6
ОБМЕН БЕЛКОВ
План
1. Гидролиз белков в органах пищеварительной системы, транспорт аминокислот в клетки.
2. Метаболизм аминокислот: реакции дезаминирования, переаминирования, декарбоксилирования.
3. Образование цистеина, метионина и др.
6. Биосинтез белка: генетический код, репликация ДНК, транскрипция, трансляция. Мутации генов. Генная инженерия.
7. Образование конечных продуктов белкового обмена: мочевины, мочевой кислоты, креатинина и др.
8. Патология белкового обмена.
Метаболизм аминокислот
Реакции переаминирования (трансаминирования)- обмен аминогруппы аминокислоты на карбонильную группу кетокислоты. Образуется новая аминокислота и новая кетокислота. Реакции обратимые. Небелковой частью ферментов их является пиридоксальфосфат (ПФ). Название фермента включает название аминокислоты, донора аминогруппы.
СООН СООН
1 1
СН2 CН3 СН2 СН3
1 1 1 1
СН2 + С=О Аланинамино - СН2 + СН-NH2
1 1 трансфераз а 1 1
СН-NH2 СООН Пиридоксаль - С=О CООН
1 фосфат 1
COOH СООН
Глутамино- Пировиноград- α-кетоглутаро- Аланин
вая кислота ная кислота вая кислота
Реакции являются основным источником в образовании одних заменимых аминокислот из других. Например, если в организме избыток аланина, но не хватает аспарагиновой кислоты, то протекает двухстадийный процесс:
Аланин + α-кетоглутарат Аланинаминотрансфераза Глутамат + Пируват;
ПФ
Глутамат + оксалоацетат Аспартатаминотрансфераза α-кетоглутарат + аспартат.
ПФ
Реакции декарбоксилирования – отщепление углекислого газа от карбоксильной группы аминокислоты. Коферментом декарбоксилаз аминокислот также является пиридоксальфосфат (ПФ). В этих реакциях образуются некоторые гормоны и нейромедиаторы, передающие импульсы в нервных клетках.
СООН СООН
1 1
СН2 СН2
1 -СО2 1
СН2 Глутаматде- СН2
1 карбоксилаза 1
СН-NH2 Пиридоксаль - СН2
1 фосфат 1
COOH NH2
Глутамино- γ-аминомасля-
вая кислота ная кислота
Реакции дезаминирования – отщепление от аминокислоты аминогруппы.
У микроорганизмов протекают реакции восстановительного, гидролитического и внутримолекулярного дезаминирования, а в клетках млекопитающих – реакции окислительного дезаминирования.
Лекция №7
ОБМЕН БЕЛКОВ (продолжение)
План
1. Генетический код.
2. Репликация ДНК, транскрипция, трансляция.
3. Мутации генов. Генная инженерия.
Биосинтез белка
В организме существует около 5 миллионов белков. Аминокислотный состав каждого из них, а у белков, имеющих четвертичную структуру, и состав их субъединиц, закодирован в гене ДНК. Место аминокислоты в будущем белке кодируется триплетом (кодоном), состоящим из трех нуклеотидов. Имеется 4 основные разновидности нуклеотидов: дезоксиаденозинмонофосфат (дАМФ), дезоксигуанозинмонофосфат, (дГМФ), дезоксицитидинмонофосфат (дЦМФ) и дезокситимидинмонофосфат (дТМФ). Последний триплет (терминирующий) не кодирует аминокислоту. Он дает сигнал об окончании сборки на поверхности расплетенной молекулы ДНК информационной РНК (иРНК), кодирующей один белок. Одну аминокислоту могут кодировать1, 2, реже - 3 или 4 триплета. Синтезу ДНК, РНК и белков в рибосомах предшествует биосинтез пуриновых или пиримидиновых нуклеотидов.
Биосинтез пуриновых нуклеотидов.
Глюкоза пентозный рибозо-5 →5-фосфорибо- + глутамин амидофосфорибо-
цикл фосфат зилдифосфат зилтрансфераза
Фосфорибо- 8 р е а к ц и й ИМФ 2 реакции АМФ
Зиламин + НАДФ-Н2,, производные фолиевой к-та ГМФ
Регуляция биосинтеза белков
Лекция №8
ОБМЕН БЕЛКОВ (продолжение)
План
1. Образование конечных продуктов белкового обмена: мочевины, мочевой кислоты, креатинина и др.
2. Патология белкового обмена
Образование конечных продуктов белкового обмена.
Лекция №9
ОБМЕН УГЛЕВОДОВ
План
- Структура и биологическая роль углеводов.
- Гидролиз углеводов в органах пищеварительной системы.
- Синтез и распад гликогена в печени.
- Обмен углеводов в тканях: анаэробный, аэробный гликолиз., глюконеогенез, пентозный цикл.
- Регуляция и патология углеводного обмена.
Окисление глюкозы в тканях
Этот процесс может протекать в отсутствии кислорода (анаэробный гликолиз), когда глюкоза расщепляется до молочной кислоты. В присутствии кислорода (аэробный гликолиз) она окисляется до углекислого газа и воды. Оба процесса, реакции которых совпадают до стадии образования пировиноградной кислоты, необходимы главным образом для генерации АТФ.
Анаэробный гликолиз – источник АТФ у анаэробных микроорганизмов. У аэробных организмов функционирует наряду с аэробным гликолизом, дополняя его.


Анаэробное окисление гликогена в мышцах. При усиленном сокращении мышц в миоциты не успевает поступать кислород. В этих условия гликоген, содержащийся в мышцах, расщепляется гликогенфосфорилазой до глюкозо-1-фосфата. Последний превращается в результате фосфоглюкомутазной реакции в глюкозо-6-фосфат, который включается в реакции анаэробного гликолиза. При этом при окислении одного остатка глюкозы генерируется в конечном итоге 3 молекулы АТФ (при образовании глюкозо-1-фосфата АТФ не затрачивается).
Физиологическая роль анаэробного гликолиза.
Этот процесс является источником АТФ в клетках, особенно при недостатке кислорода в тканях. При окислении одной молекулы глюкозы в реакциях субстратного фосфорилирования образуется 4 молекулы АТФ: по 2 молекулы его в фосфоглицераткиназной и пируваткиназной реакциях. 2 молекулы АТФ затрачиваются на начальных этапах гликолиза: в гексокиназной фосфофруктокиназной реакциях. В конечном итоге, в реакциях анаэробного гликолиза из одной молекулы глюкозы генерируется 2 молекулы АТФ. При этом образуются 2 молекулы молочной кислоты, способные закислять ткани. Поэтому молочная кислота в реакциях глюкоконеогенеза обратно превращается в глюкозу.
Лекция №10
ОБМЕН УГЛЕВОДОВ
План
- Аэробный гликолиз.
- Глюконеогенез.
- Пентозный цикл.
- Регуляция и патология углеводного обмена.
Аэробный гликолиз – основной источник АТФ в клетках. До стадии образования 3-фосфоглицеральдегида реакции его совпадают с реакциями анаэробного гликолиза. В дальнейшем, в процессе превращения двух молекул 3-фосфоглицеральдегида в две молекулы 1,3-бисфосфоглицериновой кислоты из двух молекул НАД генерируются две молекулы НАД-Н2. При окислении последних в дыхательной цепи митохондрий вырабатывается 6 молекул АТФ (2Х3). Реакции окисления двух молекул1,3-бисфосфоглицериновой кислоты до двух молекул пировиноградной кислоты также совпадают с соответствующими реакциями анаэробного гликолиза. В дальнейшем все идет по-другому: 2 молекулы пировиноградной кислоты превращаются в 2 молекулы ацетил-КоА:

При этом происходит восстановление двух молекул НАД. При окислении двух молекул НАД-Н2 в дыхательной цепи митохондрий вырабатывается 6 молекул АТФ. Реакцию катализирует пируватдегидрогеназный комплекс, отщепляющий от молекул пирувата углекислый газ (окислительное декарбоксилирование) и присоединяющий к месту отщепления –S-КоА. В состав его входят 5 коферментов: НАД, ФАД, ТиаминДФ, HS-КоА и липоевая кислота. Образующиеся 2 молекула ацетил-КоА в дальнейшем окисляются в цикле Кребса. При этом генерируются 2 молекулы ГТФ, восстанавливаются 6 (2Х3)молекул НАД и 2 молекулы ФАД. При дальнейшем окислении НАД-Н2 и ФАД-Н2 в дыхательной цепи митохондрий генерируются 22 молекулы АТФ.
Физиологическая роль аэробного гликолиза
1.Этот процесс является основным источником АТФ в клетках. При окислении одной молекулы глюкозы затрачивается 2 молекулы АТФ (в гексокиназной и фосфофруктокиназной реакциях), но генерируется 40 его молекул в реакциях
3-фосфоглицеральдегиддегидрогеназной – 6 АТФ,
фосфоглицераткиназной– 2 АТФ,
пируваткиназной – 2 АТФ,
пируватдегидрогеназной – 6 АТФ,
изоцитратдегидрогеназной – 6 АТФ,
α-кетоглутаратдегидрогеназной – 6 АТФ,
сукцинилтиокиназной – 2 АТФ,
малатдегидрогеназной– 6 АТФ.
Итого - 40 молекул АТФ.
С учетом двух затраченных молекул АТФ энергетический эффект окисления одной молекулы глюкозы в реакция аэробного окисления глюкозы составляет 38 молекул АТФ. Конечными продуктами этого процесса являются углекислый газ и вода.
2. В реакциях аэробного гликолиза образуются метаболиты, необходимые для биосинтеза других веществ. Пировиноградная, щавелевоуксусная и α-кетоглутаровая кислоты участвуют в реакциях переаминирования аминокислот. 3-фосфоглицеральдегид используется для биосинтеза глицерола, а ацетил-КоА – жирных кислот, холестерола и т.д. Из пировиноградной кислоты синтезируется щавелевоуксусная кислота, необходимая для функционирования цикла Кребса.
Глюконеогенез – образование глюкозы из неуглеводных источников: некоторых аминокислот, глицерола, молочной и пировиноградной кислот. Протекает в печени и в меньшей степени – в почках и слизистой оболочке кишечника. Является в основном процессом, обратным процессу гликолиза, реакции которого катализируют в обратном направлении те же ферменты, что и ферменты гликолиза, за исключение трех ферментов. Пируваткиназная, фосфофруктокиназная и гексоконазная реакции протекают только а одном направлении. Реакцию, обратную пируваткиназной, катализируют 2 фермента: пируваткарбоксилаза и фосфоенолпируваткарбоксикиназа. Гидролиз фруктозо-1,6-бисфосфата до фруктозо-6-фосфата катализирует фруктозо-6-фосфатаза, а расщепление глюкозо-6-фосфата до глюкозы - глюкозо-6-фосфатаза. Биосинтез четырех специфических ферментов глюконеогенеза усиливают гормоны коркового слоя надпочечников – глюкокортикоиды.
- Обратными пируваткиназной являются реакции, катализируемые
пируваткарбоксилазой (ПК-ой) и фосфоенолпируваткарбоксикиназой (ФЕПКК-ой):
СООН
СН3 СН2 CН2
1 1 ||
СО2 + С=О ПК-за С=О ФЕПКК-за O~O-РО3Н2
1 Биотин 1 -СО2 1
СООН СООН СООН
АТФ АДФ+ ГТФ ГДФ
Н3РО4
Пировиноград- Щавелевоуксус- Фосфоенолпи-
ная кислота ная кислота ровиноградная
кислота
- Обратной фосфофрукиокиназной являются реакция, катализируемая
фруктозо-1,6- бисфосфатазой (Ф-1,6-БФ-ой):
- Обратная гексокиназной является реакция, катализируемая глюкозо-6-
Фосфатазой (Г-6-Ф-ой):
Биологическая роль глюконеогенеза.
Глюконеогенез из молочной кислоты усиливается при повышении в тканях ее уровня, вызванном физическими нагрузками или недостатком в организме кислорода. При этом молочная кислота вначале превращается в пировиноградную в результате реакции, катализируемой лактатдегидрогеназой. Глюконеогенез из аминокислот усиливается при хронических стрессовых ситуациях, сахарном диабете и других состояниях, приводящих к недостатку в организме углеводов. При этом некоторые аминокислоты предварител






