Пищевые жиры: нормы суточного потребления, переваривание и всасывание у детей. Ресинтез жиров в клетках кишечника. Хиломикроны, ЛПОНП. Факторы просветления хилезной плазмы крови. Роль желчных кислот в переваривании и всасывании жиров.
Нормы суточного потребления: потребность в липидах взрослого организма составляет 80-100 г в сутки, из них растительных жиров должно быть не менее 30%. С пищей в основном поступают триацилглицеролы, фосфолипиды и эфиры ХС.
Особенности переваривания жира у детей: У младенцев клетками слизистой корня языка и глотки (железы Эбнера) при сосании секретируется лингвальная липаза, продолжающая свое действие и в желудке.
У грудных младенцев и детей младшего возраста липаза желудка более активна, чем у взрослых, так как кислотность в желудке детей около 5,0. Помогает и то, что жиры молока эмульгированы. Жиры у младенцев дополнительно перевариваются за счет липазы женского молока, в коровьем молоке липаза отсутствует. Благодаря таким преимуществам у детей грудного возраста в желудке происходит 25-50% всего липолиза.
В двенадцатиперстной кишке гидролиз жира дополнительно осуществляется панкреатической липазой. До 7 лет активность панкреатической липазы невысока, что ограничивает способности ребенка к перевариванию пищевого жира, ее активность достигает максимума только к 8-9 годам. Но, тем не менее, это не мешает ребенку уже в первые месяцы жизни гидролизовать почти 100% пищевого жира и иметь 95% всасывания.В грудном возрасте содержание желчных кислот в желчи постепенно увеличивается примерно в три раза, позднее этот рост замедляется
Всасывание липидов
После расщепления полимерных липидных молекул полученные мономеры всасываются в верхнем отделе тонкого кишечника в начальные 100 см. В норме всасывается 98% пищевых липидов.
1. Короткие жирные кислоты (не более 10 атомов углерода) всасываются и переходят в кровь без каких-либо особенных механизмов. Этот процесс важен для грудных детей, т.к. молоко содержит в основном коротко- и среднецепочечные жирные кислоты. Глицерол тоже всасывается напрямую.

2. Другие продукты переваривания образуют с желчными кислотами мицеллы с гидрофильной поверхностью и гидрофобным ядром. Далее в энтероците происходит распад мицеллы на триацилглицеролы, фосфоглицериды, холестеролы и белок апо-В48, которые в составе хиломикронов 90% попадают в лимфу (через грудной проток)- а затем в кровь.
Хиломикроны. ЛПОНП.
Хиломикроны. Общая характеристика
+формируются в кишечнике из ресинтезированных жиров,
+являются самыми крупными липопротеинами, их размер от 100 до 1200 нм (0,1-1,2 мкм)
+в их составе преобладают ТАГ, мало белка, фосфолипидов и холестерола (2% белка, 87% ТАГ, 2% ХС, 5% эфиров ХС, 4% фосфолипидов),
+ основным апобелком является апоВ-48, это структурный липопротеин, в плазме крови получают от ЛПВП белки апоС-II и апоЕ
+в норме натощак не обнаруживаются, в крови появляются после приема пищи, поступая из лимфы через грудной лимфатический проток, и полностью исчезают через 10-12 часов,
+не атерогенны.
Функция: Транспорт экзогенных ТАГ из кишечника в ткани, запасающие или использующие жирные кислоты, в основном в жировую ткань, миокард, скелетные мышцы и лактирующую молочную железу, в меньшей степени в легкие, костный мозг, почки, селезенку. На эндотелии капилляров этих тканей имеется фермент липопротеинлипаза.
ЛПОНП.Липопротеины очень низкой плотности :
+синтезируются в печени из эндогенных и экзогенных липидов,
+в их составе преобладают ТАГ, около 40% от массы составляют белок, фосфолипиды и холестерол (8% белка, 60% ТАГ, 6% ХС, 12% эфиров ХС, 14% фосфолипидов),
+основным белком является апоВ-100, выполняющий структурную функцию,
+в норме концентрация 1,3-2,0 г/л,
+слабо атерогенны.
Функция
Транспорт эндогенных и экзогенных ТАГ от печени в ткани, запасающие и использующие жирные кислоты, т.е. в те же ткани, что и хиломикроны.
Тканевой распад и синтез триглицеридов. Особенности синтеза триглицеридов в жировой клетке. Регуляция липогенеза и липолиза, ключевые ферменты. Особенности обмена липидов в детском возрасте.
Окисление высших жирных кислот. Этапы окисления. Роль карнитина. Энергетическая характеристика окисления. Особенности окисления ненасыщенных жирных кислот и кислот с разветвленной структурой. Бурая жировая ткань, особенности ее структуры и функции.
Этапы окисления
1. Прежде, чем проникнуть вматрикс митохондрий и окислиться, жирная кислота должна активироваться в цитозоле. Это осуществляется присоединением к ней коэнзима А с образованием ацил-SКоА. Ацил-SКоА является высокоэнергетическим соединением. Необратимость реакции достигается гидролизом дифосфата на две молекулы фосфорной кислоты.
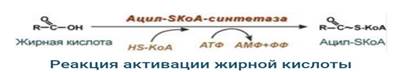
2. Ацил-SКоА не способен проходить через митохондриальную мембрану, поэтому существует способ переноса жирной кислоты в комплексе с витаминоподобным веществом карнитином (витамин В11). На наружной мембране митохондрий имеется фермент карнитин-ацилтрансфераза I.
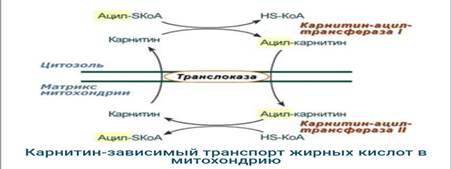
3. После связывания с карнитином жирная кислота переносится через внутреннюю митохондриальную мембрану транслоказой. На внутренней стороне этой мембраны фермент карнитин-ацилтрансфераза II вновь образует ацил-SКоА, который вступает на путь β-окисления.
4. Процесс собственно β-окисления состоит из 4-х реакций, повторяющихся циклически. В них последовательно происходит окисление (ацил-SКоА-дегидрогеназа), гидратирование (еноил-SКоА-гидратаза) и вновь окисление 3-го атома углерода (гидроксиацил-SКоА-дегидрогеназа). В последней, трансферазной, реакции от жирной кислоты отщепляется ацетил-SКоА. К оставшейся (укороченной на два углерода) жирной кислоте присоединяется HS-КоА, и она возвращается к первой реакции. Все повторяется до тех пор, пока в последнем цикле не образуются два ацетил-SКоА.

Строение
Клетки белой и бурой жировой ткани значительно отличаются друг от друга. Клетка бурой жировой ткани имеет по всей цитоплазме много мелких жировых капель, окружающих большое количество митохондрий, обильно снабжена окончаниями симпатических нервов и кровеносными сосудами. В эмбриогенезе она развивается из миобластов миотома сомитов, т.е. из предшественников скелетной мускулатуры, а не из предшественников соединительной ткани, подобно белым адипоцитам.
Бурая жировая ткань у младенцев составляет около 5% от массы их тела. Она расположена подкожно, между лопатками, в виде небольших отложений вокруг мышц и кровеносных сосудов шеи, в щеках (комочки Биша), а также в большом количестве – в подмышечных впадинах. Более глубокие отложения находятся в переднем средостении, вдоль позвоночника, вокруг почек, надпочечников, аорты, между лопатками, около почек и щитовидной железы
Функции
Функцией бурой жировой ткани является оперативное обеспечение организма тепловой энергией (адаптивныйтермогенез). Основной механизм термогенеза определяется тем, что в митохондриях клеток бурых адипоцитов процессы окисления и фосфорилирования не сопряжены (разобщены) и при окислении поступающих сюда жирных кислот происходит, в основном, выделение тепла, а не синтез АТФ.
Разобщение окисления и фосфорилирования обеспечивают специальные белки – термогенины (разобщающие белки, uncouplingproteins, UCP).
Адаптивный термогенез – это повышение теплопродукции в ответ:
- на изменение окружающей среды (снижение температуры) и на переохлаждение организма. При этом активируется симпатическая нервная система и стимулируется липолиз в бурыхадипоцитах,
- на стимуляцию катехоламинами (через β3-адренорецепторы) – повышение транскрипции гена UCP-1, что приводит к увеличению энергозатрат,
- на избыток потребления пищи или изменение ее состава (поступление жиров),
- на секрецию лептина (усиление транскрипции гена UCP-1),
- на какие-то патологические воздействия.
Механизм биосинтеза.
Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо декарбоксилируется до ацетона. После синтеза все три соединения поступают в кровь и разносятся по тканям. Ацетон, как летучее вещество, легко удаляется с выдыхаемым воздухом и потом. Все кетоновые тела могут выделяться с мочой.
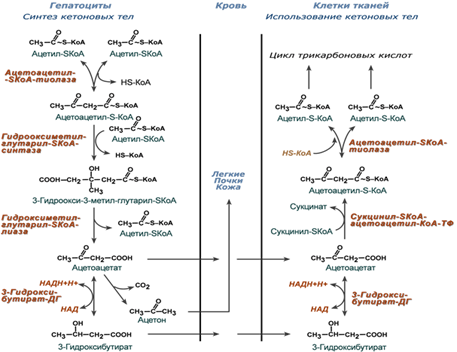
Биологическая роль
Энергетическая - являются альтернативным глюкозе источником энергии:
+ в период голодания мышцами, почками, кишечником, мозгом и нервной тканью.
+при длительной физической работе мышечной тканью.
+при сахарном диабете первого типа (инсулинзависимом).вгепатоците глюкозы может быть много, т.к. глюкоза проникает в него через ГлюТ-2 без участия инсулина. Однако, точно также как при голодании, соотношение инсулин/глюкагон низкое, и оксалоацетат очень активно уходит на синтез глюкозы, что тормозит ЦТК и отправляет ацетил-SKoA на образование кетоновых тел.
Окисление кетоновых тел- аэробный процесс, происходящий в матриксе митохондрий (при окислении ацетоацетата выход энергии равен 23 АТФ на 1 моль субстрата и 26 молям АТФ на 1 моль бетта-гидроксибутирата)
Биосинтез жирных кислот.
Биосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покоя или после еды.
Условно можно выделить 4 этапа биосинтеза:
1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот.
2. Перенос ацетил-SКоА из митохондрий в цитозоль:
- может быть в комплексе с карнитином, подобно тому как переносятся внутрь митохондрии высшие жирные кислоты, но здесь транспорт идет в другом направлении,
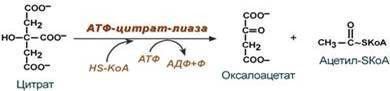
- обычно в составе лимонной кислоты, образующейся в первой реакции ЦТК.
Поступающий из митохондрий цитрат в цитозоле расщепляется АТФ-цитрат-лиазой до оксалоацетата и ацетил-SКоА.
3. Образование малонил-SКоА из ацетил-SКоА.
Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов.

4. Синтез пальмитиновой кислоты.
Осуществляется мультиферментным комплексом " синтаза жирных кислот " (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ).
Ацил-переносящий белок включает производное пантотеновой кислоты – 6-фосфопантетеин (ФП), имеющий HS-группу, подобно HS-КоА. Один их ферментов комплекса, 3-кетоацил-синтаза, также имеет HS-группу в составе цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтеза необходим НАДФН.

4.Удлинение цепи жирных кислот
Синтезированная пальмитиновая кислота при необходимости поступает в эндоплазматическийретикулум. Здесь с участием малонил-S-КоА и НАДФН цепь удлиняется до С18 или С20. Присоединение углеродов идет по карбоксильной группе (С1).
Биосинтез холестерина
Синтез холестерола в организме составляет примерно 0,5-0,8 г/сут, примерно 50% образуется в печени, около 15% в кишечнике. Все клетки организма способны синтезировать холестерол.
Биосинтез холестерола происходит в эндоплазматическомретикулуме. Источником всех атомов углерода в молекуле является ацетил-SКоА, поступающий сюда из митохондрий в составе цитрата, также как при синтезе жирных кислот. При биосинтезе холестерола затрачивается 18 молекул АТФ и 13 молекул НАДФН.
Образование холестерола идет более чем в 30 реакциях, которые можно сгруппировать в несколько этапов.
1. Синтез мевалоновой кислоты.
Первые две реакции синтеза совпадают с реакциями кетогенеза, но после синтеза 3-гидрокси-3-метилглутарил-SКоА вступает в действие фермент гидроксиметил-глутарил-SКоА-редуктаза (ГМГ-SКоА-редуктаза), образующая мевалоновую кислоту.
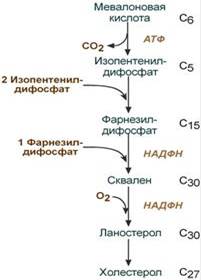
2. Синтез изопентенилдифосфата. На этом этапе при использовании АТФ мевалоновая кислота трижды фосфорилируется. Затем промежуточный продукт декарбоксилируется и дефосфорилируется с получением изопентенилдифосфата.
3. После объединения трех молекул изопентенилдифосфата (если не считать промежуточных превращений) синтезируется фарнезилдифосфат.
4. Синтез сквалена происходит присвязывания двух остатков фарнезилдифосфата.
5. После сложных реакций линейныйскваленциклизуется в ланостерол.
6. Удаление лишних метильных групп, восстановление и изомеризация молекулы приводит к появлению холестерола.
Этерификация холестерина в плазме крови и клетках (ЛХАТ, АХАТ). Особенности метаболизма холестерина в коже, половых железах, коре надпочечников и печени. Синтез желчных кислот. Структура первичных и вторичных желчных кислот. Парные желчные кислоты. Выведение холестерина из организма. Биологическая роль и кругооборот желчных кислот.
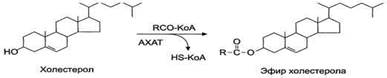
Этерификация холестерина- это образование эфиров холестерина.
Внутри ЛПВП активно протекает реакция при участии лецитин:холестерол-ацилтрансферазы (ЛХАТ-реакция). В этой реакции остаток полиненасыщенной жирной кислоты переносится от 2-го положения фосфатидилхолина (из оболочки самого ЛПВП) на получаемый свободный холестерин с образованием лизофосфатидилхолина (лизоФХ) и эфиров ХС. ЛизоФХ остается внутри ЛПВП, эфир холестерина отправляется в ЛПНП.

В стенке кишечника основная часть холестерина этерифицируется под влиянием фермента- ацил-СоА-холестеринацилтрансферазы (АХАТ).
Синтез фосфатидной кислоты
Жирные кислоты, поступающие из крови при распаде ХМ, ЛПОНП или синтезированные в клетке из глюкозы, также должны активироваться. Они превращаются в ацил-SКоА в АТФ-зависимой реакции.

При наличии глицерол-3-фосфата и ацил-SКоА синтезируется фосфатидная кислота.

Липотропные факторы- это природные вещества или лекарственные препараты, способствующие синтезу фосфолипидов и препятствующие отложению триацилглицеролов в печени.
· Холин (самый распространенный аминоспирт в фосфолипидах)
· Метионин (незаменимая аминокислота, предшественник S-аденозилметионина, универсальный донор метильной группы)
· Фолиевая кислота и витамин В12
· Полиненасыщенные жирные кислоты и другие соединения.
Пищевые жиры: нормы суточного потребления, переваривание и всасывание у детей. Ресинтез жиров в клетках кишечника. Хиломикроны, ЛПОНП. Факторы просветления хилезной плазмы крови. Роль желчных кислот в переваривании и всасывании жиров.
Нормы суточного потребления: потребность в липидах взрослого организма составляет 80-100 г в сутки, из них растительных жиров должно быть не менее 30%. С пищей в основном поступают триацилглицеролы, фосфолипиды и эфиры ХС.
Особенности переваривания жира у детей: У младенцев клетками слизистой корня языка и глотки (железы Эбнера) при сосании секретируется лингвальная липаза, продолжающая свое действие и в желудке.
У грудных младенцев и детей младшего возраста липаза желудка более активна, чем у взрослых, так как кислотность в желудке детей около 5,0. Помогает и то, что жиры молока эмульгированы. Жиры у младенцев дополнительно перевариваются за счет липазы женского молока, в коровьем молоке липаза отсутствует. Благодаря таким преимуществам у детей грудного возраста в желудке происходит 25-50% всего липолиза.
В двенадцатиперстной кишке гидролиз жира дополнительно осуществляется панкреатической липазой. До 7 лет активность панкреатической липазы невысока, что ограничивает способности ребенка к перевариванию пищевого жира, ее активность достигает максимума только к 8-9 годам. Но, тем не менее, это не мешает ребенку уже в первые месяцы жизни гидролизовать почти 100% пищевого жира и иметь 95% всасывания.В грудном возрасте содержание желчных кислот в желчи постепенно увеличивается примерно в три раза, позднее этот рост замедляется
Всасывание липидов
После расщепления полимерных липидных молекул полученные мономеры всасываются в верхнем отделе тонкого кишечника в начальные 100 см. В норме всасывается 98% пищевых липидов.
1. Короткие жирные кислоты (не более 10 атомов углерода) всасываются и переходят в кровь без каких-либо особенных механизмов. Этот процесс важен для грудных детей, т.к. молоко содержит в основном коротко- и среднецепочечные жирные кислоты. Глицерол тоже всасывается напрямую.
2. Другие продукты переваривания образуют с желчными кислотами мицеллы с гидрофильной поверхностью и гидрофобным ядром. Далее в энтероците происходит распад мицеллы на триацилглицеролы, фосфоглицериды, холестеролы и белок апо-В48, которые в составе хиломикронов 90% попадают в лимфу (через грудной проток)- а затем в кровь.
Ресинтез жиров в клетках кишечника.
Ресинтез липидов – это синтез липидов в стенке кишечника из поступающих сюда экзогенных жиров, одновременно могут использоваться и эндогенные жирные кислоты, поэтому ресинтезированные жиры отличаются от пищевых и более близки по составу к "своим" жирам. Основная задача этого процесса – связать поступившие с пищей средне- и длинноцепочечные жирные кислоты со спиртом – глицеролом или холестеролом. Это, во-первых, ликвидирует их детергентное действие на мембраны и, во-вторых, создает их транспортные формы для переноса по крови в ткани.
1) Активация жирной кислоты
Поступившая в энтероцит (как и в любую другую клетку) жирная кислота обязательно активируется через присоединение коэнзима А. Образовавшийся ацил-SКоА участвует в реакциях синтеза эфиров холестерола, триацилглицеролов и фосфолипидов

2) Ресинтез эфиров холестерола
Холестеролэтерифицируется с использованием ацил-SКоА и фермента ацил-SКоА:холестерол-ацилтрансферазы (АХАТ).

3) Ресинтез ТАГ (триацилглицерола): Для ресинтеза ТАГ есть два пути:
Первый путь, основной – 2-моноацилглицеридный – происходит при участии экзогенных 2-МАГ и ЖК в гладком эндоплазматическом ретикулумеэнтероцитов: мультиферментный комплекс триацилглицерол-синтазы формирует ТАГ.
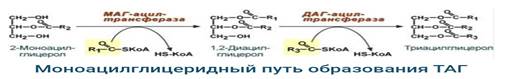
существует второй, глицеролфосфатный, путь в шероховатом эндоплазматическом ретикулуме. Источником глицерол-3-фосфата служит окисление глюкозы. Здесь можно выделить следующие реакции:
+Образование глицерол-3-фосфата из глюкозы.
+Превращение глицерол-3-фосфата в фосфатидную кислоту.
+Превращение фосфатидной кислоты в 1,2-ДАГ.
+Синтез ТАГ.

Хиломикроны. ЛПОНП.
Хиломикроны. Общая характеристика
+формируются в кишечнике из ресинтезированных жиров,
+являются самыми крупными липопротеинами, их размер от 100 до 1200 нм (0,1-1,2 мкм)
+в их составе преобладают ТАГ, мало белка, фосфолипидов и холестерола (2% белка, 87% ТАГ, 2% ХС, 5% эфиров ХС, 4% фосфолипидов),
+ основным апобелком является апоВ-48, это структурный липопротеин, в плазме крови получают от ЛПВП белки апоС-II и апоЕ
+в норме натощак не обнаруживаются, в крови появляются после приема пищи, поступая из лимфы через грудной лимфатический проток, и полностью исчезают через 10-12 часов,
+не атерогенны.
Функция: Транспорт экзогенных ТАГ из кишечника в ткани, запасающие или использующие жирные кислоты, в основном в жировую ткань, миокард, скелетные мышцы и лактирующую молочную железу, в меньшей степени в легкие, костный мозг, почки, селезенку. На эндотелии капилляров этих тканей имеется фермент липопротеинлипаза.
ЛПОНП.Липопротеины очень низкой плотности :
+синтезируются в печени из эндогенных и экзогенных липидов,
+в их составе преобладают ТАГ, около 40% от массы составляют белок, фосфолипиды и холестерол (8% белка, 60% ТАГ, 6% ХС, 12% эфиров ХС, 14% фосфолипидов),
+основным белком является апоВ-100, выполняющий структурную функцию,
+в норме концентрация 1,3-2,0 г/л,
+слабо атерогенны.
Функция
Транспорт эндогенных и экзогенных ТАГ от печени в ткани, запасающие и использующие жирные кислоты, т.е. в те же ткани, что и хиломикроны.






