| Название рефлекса
| Применяемое раздражение
| Характер рефлекторной реакции
| Локализация нейронов, участвующих в рефлексе
|
| Сухожильные проприорецептивные рефлексы:
|
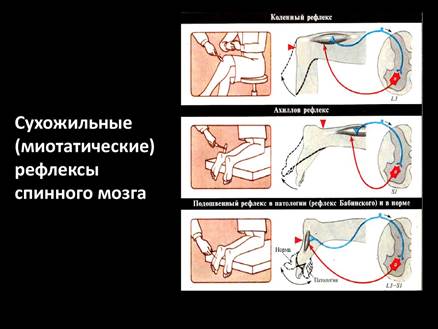
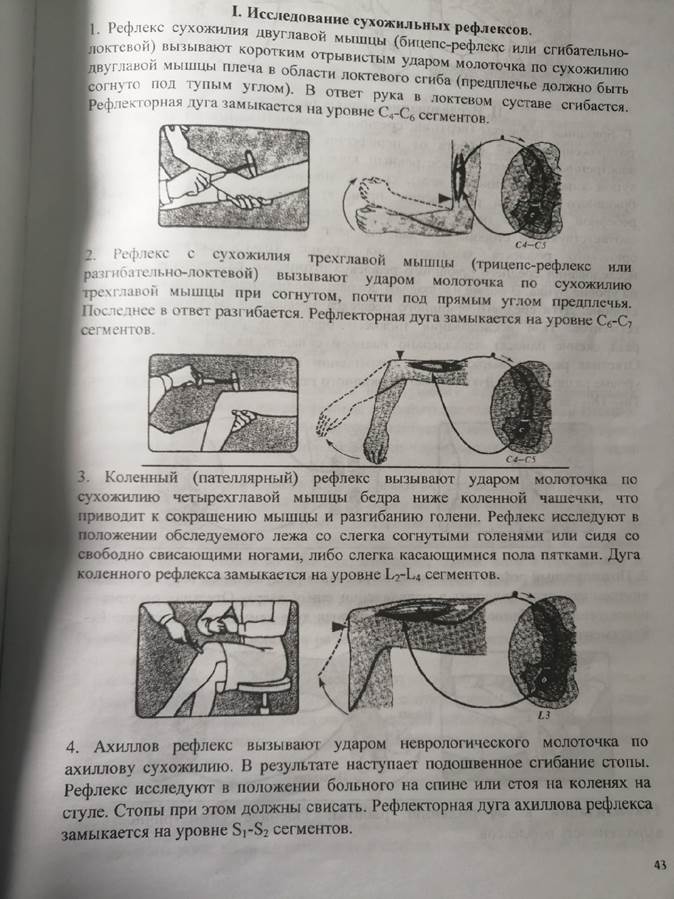
| сгибательно- локтевой (бицепс- рефлекс)
| Удар молоточком по сухожилию двуглавой мышцы плеча (рука слегка согнута в локте)
| Сокращение двуглавой мышцы плеча и сгибание руки
| V-VI шейные сегменты спинного мозга
|
| коленный
| Удар молоточком по сухожилию четырехглавой мышцы бедра ниже надколенника
| Сокращение четырехглавой мышцы бедра и разгибание голени
| II-IV поясничные
сегменты
|
| ахиллов
| Удар по ахиллову сухожилию
| Подошвенное сгибание стопы
| I-II крестцовые
сегменты
|
| Брюшные рефлексы:
| Штриховое раздражение кожи:
| Сокращение соответствующих участков брюшной мускулатуры
|
|
| - верхний
| параллельно нижним ребрам
|
| VIII-IX грудные сегменты
|
| средний
| на уровне пупка (горизонтально)
|
| IX-XII грудные сегменты
|
| нижний
| параллельно паховой
складке
|
| 1-Й поясничные сегменты
|
| Кремастерный яичковый рефлекс
| Штриховое раздражение внутренней поверхности бедра
| Сокращение мышцы, поднимающей яичко
| I-II поясничные сегменты
|
| Анальный рефлекс
| Штрих или укол вблизи заднего прохода
| Сокращение наружного сфинктера прямой кишки
| IV-V крестцовые сегменты
|
| Подошвенный рефлекс
| Слабое штриховое раздражение подошвы
| Сгибание пальцев и стопы
| I- II крестцовые сегменты
|
|
| Сильное раздражение подошвы
| Разгибание пальцев и сгибание ноги
|
|
При вызывании рефлексов необходимо добиваться от больного возможно более полного расслабления исследуемой конечности. Удары молоточком следует наносить с одинаковой силой. При оценке рефлексов обращают внимание на их выраженность и симметричность. Поэтому исследовать одни и те же рефлексы надо справа и слева и сразу их сравнивать. Выраженность рефлексов у здоровых людей может индивидуально колебаться, в частности, симметричное положение или оживление рефлексов при полной сохранности мышечной силы можно расценивать как вариант нормы.
Наблюдаются следующие изменения рефлексов: понижение или утрата (при повреждении рефлекторной дуги), повышение, и извращение (при поражении пирамидной системы и растормаживании сегментарного аппарата спинного мозга).
Сухожильные рефлексы:
Кожные рефлексы:

25. Двигательные ядра черепно-мозговых нервов, их функция.




26. Вестибулярные ядра, вестибулярный тракт и его роль в регуляции мышечного тонуса.

Вестибулярный анализатор играет важную роль в регуляции мышечного тонуса и сохранении позы организма, он обеспечивает возникновение акселерационных ощущений, т. е. при прямолинейном и вращательном ускорении движения тела, а также при изменениях положения головы.
Роль вестибулярных ядер в возбуждении антигравитационных мышц.
Антигравитационные мышцы - это набор групп мышц, основная функция которых - поддерживать силу тяжести для поддержания определенной позы у здорового человека.
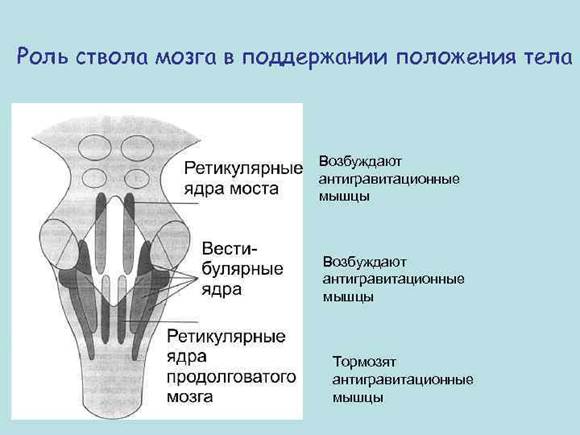
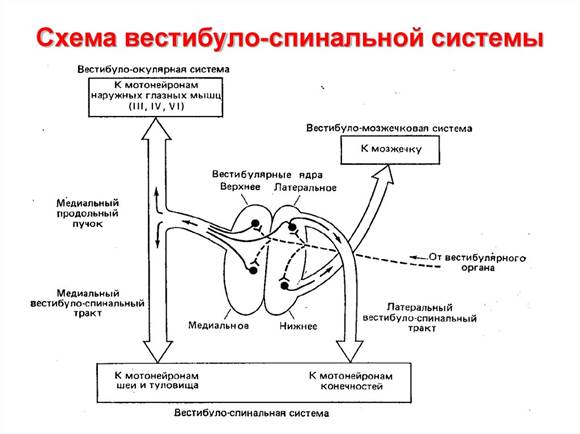
Все вестибулярные ядра функционируют вместе с ретикулярными ядрами моста для управления антигравитационными мышцами. Вестибулярные ядра посылают сильные возбуждающие сигналы к антигравитационным мышцам по латеральному и медиальному вестибулоспинальным трактам, расположенным в переднем столбе спинного мозга. Без этой поддержки вестибулярных ядер мостовая ретикулярная система потеряла бы большую часть своего возбуждающего влияния на осевые антигравитационные мышцы.
Специфическая роль вестибулярных ядер, однако, состоит в избирательной регуляции возбуждения различных антигравитационных мышц для подержания равновесия в ответ на сигналы от вестибулярного аппарата.
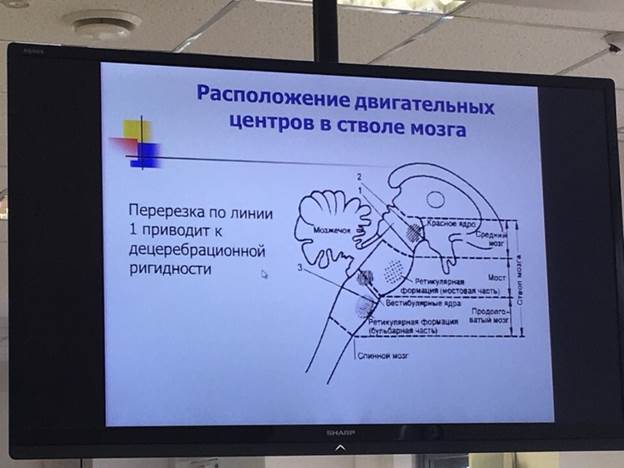
У децеребрированного животного развивается спастическая ригидность. Когда ствол мозга животного пересекают ниже уровня середины среднего мозга так, чтобы интактными остались ретикулярные системы моста и продолговатого мозга, а также вестибулярная система, развивается состояние, называемое децеребрационной ригидностью. При этом ригидность развивается не во всех мышцах тела, а в антигравитационных мышцах шеи и туловища и в разгибателях ног. Причиной децеребрационной ригидности является блокада сильного влияния коры большого мозга, красных ядер и базальных ганглиев на ретикулярные ядра продолговатого мозга. Без этого влияния тормозное действие ретикулярной формации продолговатого мозга не проявляется, и мощная гиперактивность возбуждающей системы моста ведет к развитию ригидности.
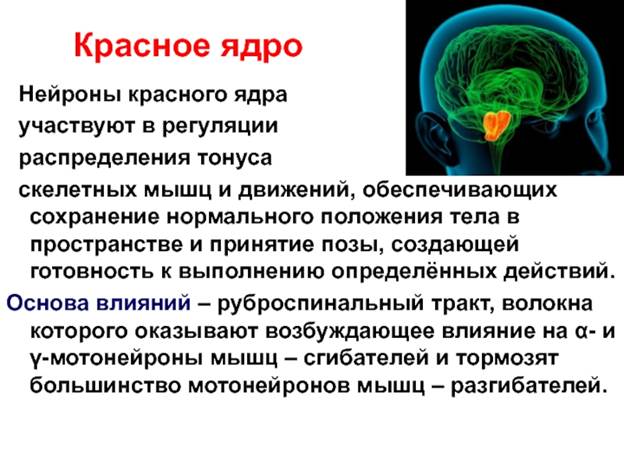
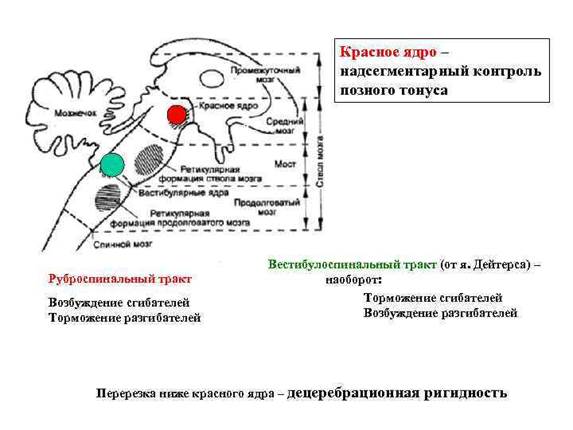
27. Красное ядро, руброспинальный тракт и его роль в регуляции мышечного тонуса. Децеребрационная ригидность
Красные ядра расположены в верхней части ножки мозга (средний мозг). К нему идут нервные пути от коры полушарий, подкорковых ядер, мозжечка. От него идет руброспинальный тракт к мотонейронам сгибателей спинного и ретикулярной формации продолговатого мозга. В связи с различным функциональным значением ядра Дейтерса и красного ядра, при перерезке ствола между средним и продолговатым мозгом у животных возникает децеребрационная ригидность – это резкое повышение тонуса всех мышц разгибателей. Голова животного запрокидывается, спина выгибается, конечности вытягиваются. Механизм децеребрационной ригидности заключается в том, что красное ядро, активируя мотонейроны сгибателей, через вставочные тормозные нейроны тормозит мотонейроны разгибателей. Одновременно исключается тормозящее влияние красного ядра на ретикулярную формацию продолговатого мозга, возле ядра Дейтерса. В отсутствии влияния красного ядра преобладает возбуждающее действие ядра Дейтерса на мотонейроны разгибателей.
Красное ядро, расположенное в среднем мозге, функционирует в тесной связи с кортикоспинальным трактом. Ядро получает большое число волокон непосредственно от первичной моторной коры через кортикорубралъный тракт, а также множество коллатералей от кортикоспинального тракта, когда он проходит через средний мозг. Эти волокна формируют синапсы в нижней, крупноклеточной (магноцеллюлярной) части красного ядра, где находятся крупные нейроны, по размерам схожие с клетками Беца в моторной коре. Эти нейроны дают начало руброспиналъному тракту, который переходит на противоположную сторону в нижней части ствола мозга и спускается в боковые столбы спинного мозга, следуя в непосредственной близости к кортикоспинальному тракту и впереди него.
Руброспинальные волокна заканчиваются в основном на вставочных нейронах промежуточных областей серого вещества спинного мозга вместе с кортикоспинальными волокнами, а некоторые из руброспинальных волокон формируют синапсы непосредственно на передних мотонейронах, как и некоторые кортикоспинальные волокна.
Красное ядро также тесно связано с мозжечком, подобно связям между мозжечком и моторной корой.
Функции кортикоруброспинальной системы. Крупноклеточная часть красного ядра имеет соматографическое представительство всех мышц тела, как и моторная кора. Следовательно, точечная стимуляция этой части красного ядра вызывает сокращение одиночной мышцы или небольшой группы мышц. Однако тонкость представительства разных мышц гораздо меньше, чем в моторной коре, особенно для человека, поскольку размеры красного ядра у людей относительно невелики. Кортикоруброспинальный путь функционирует в качестве вспомогательного пути для передачи относительно обособленных сигналов от моторной коры к спинному мозгу. Если кортикоспинальные волокна разрушаются при полной сохранности кортикоруброспинального пути, отдельные движения могут осуществляться, однако резко нарушается регуляция тонких движений пальцев рук и кистей. Движения запястья еще возможны, что не происходит в случае, если кортикоруброспинальный путь также блокируется. Следовательно, путь через красное ядро к спинному мозгу связан с кортикоспинальной системой. Более того, руброспинальный тракт лежит в боковых столбах спинного мозга вместе с кортикоспинальным трактом и заканчивается на вставочных и моторных нейронах, контролирующих дистальные мышцы конечностей. В связи с этим кортикоспинальный и руброспинальный тракты вместе называют латеральной моторной системой спинного мозга в отличие от вестибулоретикулоспинальной системы, лежащей медиально в спинном мозге, которую называют медиальной моторной системой спинного мозга.

28. Статические и статокинетические рефлексы.
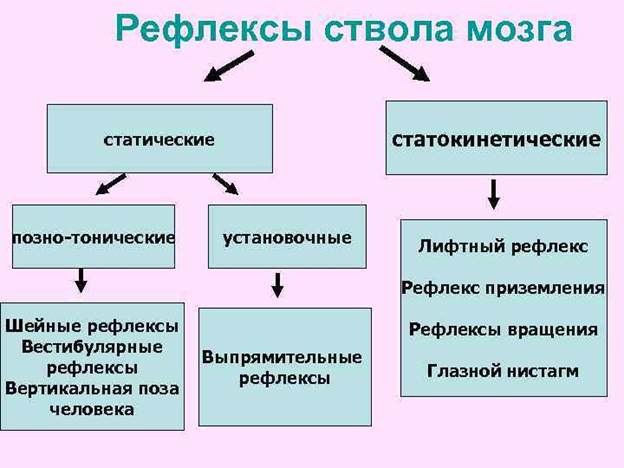
Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменении положения головы в пространстве — это так называемые лабиринтные рефлексы. возникающие в результате раздражения рецепторов вестибулярного аппарата; 2) при изменении положения головы по отношению к туловищу — шейные рефлексы, с проприорецепторов мышц шеи и 3) при нарушении нормальной позы тела — выпрямительные рефлексы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус сгибателей (лабиринтный рефлекс). Выпрямительные рефлексы — это последовательные сокращения мышц шеи и туловища, которые обеспечивают возвращение тела в вертикальное положение теменем кверху. У человека они проявляются, например, во время ныряния.
Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. Например, при быстром подъеме усиливается тонус сгибателей, и человек приседает, а при быстром спуске усиливается тонус разгибателей, и человек выпрямляется — это так называемый лифтный рефлекс. При вращении тела реакции противовращения проявляются в отклонении головы, тела и глаз в сторону, противоположную движению. Движение глаз со скоростью вращения тела, но в противоположную сторону и быстрое возвращение в исходное положение — нистагм глаз — обеспечивают сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.

Статические:


Статокинетические:

Примерами статокинетических рефлексов могут быть рефлексы лифта, компаса (противовращения) и готовности к прыжку. Рефлекс лифта проявляется увеличением тонуса разгибателей конечностей при опускании платформы, на которой находится животное, и сгибателей – при подъёме.
29. Ретикулярная формация ствола мозга и её нисходящие влияния на рефлекторную деятельность спинного мозга.
Нисходящие тормозные влияния ретикулярной формации на спинной мозг
Более ста лет назад в 1862 г. И. М. Сеченов установил факт угнетения всех спинальных рефлексов (сгибательных и разгибательных) при раздражении стволовой части мозга. Это было открытием центрального торможения и одновременно открытием ретикулоспинальной системы. Локальное электрическое раздражение гигантоклеточного ядра ретикулярной формации продолговатого мозга вызывает неспецифическое торможение сгибательных и разгибательных спинальных рефлексов. Эти неспецифические супраспинальные влияния по ретикулоспинальному тракту достигают мотонейронов спинного мозга и увеличивают порог и скрытый период их ответов на рефлекторные воздействия. Таким образом тормозятся структуры СМ.
В нисходящих путях от ретикулярной формации существуют 2 системы:
● сгибательная (латеральный ретикулоспинальный путь). Возбуждаются сгибатели, тормозятся разгибатели.
● разгибательная (медиальный ретикулоспинальный путь). Возбуждаются разгибатели, тормозятся сгибатели.
В ходе экспериментов с локальной стимуляцией ретикулярной формации выяснилось наличие зон, дающих эффект противоположной полярности, т. е. облегчающее влияние на спинно-мозговые рефлексы. Так, например, электрическое раздражение латеральных зон ретикулярной формации моста снижает порог и укорачивает скрытый период спинальных рефлексов.
Ретикулярная формация как один из двигательных центров стволовой части мозга может выступать не только в роли регулятора возбудимости спинальных мотонейронов, но и принимать участие в процессах, связанных с поддержанием позы и организацией целенаправленных движений.
Восходящие активирующие влияния ретикулярной формации на головной мозг
Если через хронически вживленные электроды раздражать центральные части ретикулярной формации ствола, то кошка, находящаяся в сонном состоянии, пробуждается и у нее появляется ориентировочная реакция. Эта поведенческая реакция пробуждения сопровождается характерными изменениями частотного спектра электроэнцефалограммы, переходом от регулярных, высоковольтных колебаний альфа-ритма к низковольтным колебаниям бета-ритма. Данная электроэнцефалографическая реакция получила название реакции десинхронизации. Она имеет генерализованный характер и регистрируется от обширных областей коры головного мозга.
В остром опыте перерезка стволовой части мозга на уровне среднего мозга и, таким образом, разрушение восходящих путей от ретикулярной формации ствола переводят животное в сноподобное коматозное состояние (спящий мозг, по Бремеру) с соответствующими изменениями характера электроэнцефалограммы.
В состав ретикулярной формации мозгового ствола входят не только структуры, при возбуждении которых животное просыпается и становится активным (настораживание, принюхивание и пр.), но и структуры, активация которых вызывает засыпание животного. Это ядра шва. В окончаниях своих аксонов (синапсах) нейроны этих ядер выделяют серотонин. В опытах на животных было показано, что локальное разрушение этих ядер приводит к хронической бессоннице животного, которая может заканчиваться смертью. Известно, что истощение серотонина мозга у человека также приводит к хронической бессоннице.
Приведенные выше экспериментальные факты послужили основанием для заключения, что ретикулярная формация является структурой, отвечающей за состояние бодрствования, структурой, формирующей восходящую активирующую ретикулярную систему, которая поддерживает на определенном уровне возбудимость промежуточного мозга и коры больших полушарий. Важнейшая функция восходящих путей РФ – регуляция цикла «сон-бодрстование» и регуляция уровня сознания. Согласно современным представлениям, переход коры к активному состоянию связан с колебаниями количества восходящих сигналов от ретикулярной формации ствола. Количество этих сигналов зависит от поступления в ретикулярную формацию сенсорных импульсов по коллатералям специфических афферентных восходящих путей.
Практически к ретикулярной формации приходит вся информация от всех органов чувств по коллатералям от спиноретикулярного тракта, проприоспинальных путей, афферентных черепно-мозговых нервов, от таламуса и гипоталамуса, от моторных и сенсорных областей коры.






