МЕТАБОЛИЗМ УГЛЕВОДОВ И ЕГО РЕГУЛЯЦИЯ
Углеводы – класс органических веществ, включающий в себя альдегиды и кетоны многоатомных спиртов, а также их производные. В природе углеводы существуют в форме моносахаридов, дисахаридов (в их составе – два моносахаридных фрагмента), олигосахаридов (содержат от трех до десяти моносахаридных фрагментов) и полисахаридов (содержат от десятков до десятков тысяч моносахаридных фрагментов).
Углеводы выполняют в организме несколько важных функций.
ФУНКЦИИ УГЛЕВОДОВ В ОРГАНИЗМЕ ЧЕЛОВЕКА
1. ЭНЕРГЕТИЧЕСКАЯ. В обычных условиях именно углеводы при их окислении являются главным источником энергии для большинства тканей организма человека, а для некоторых из них – и единственным.
2. СТРУКТУРНАЯ (ПЛАСТИЧЕСКАЯ).
А) Моносахариды рибоза и дезоксирибоза входят в состав мононуклеотидов, из которых построены нуклеиновые кислоты (ДНК и РНК).
Б) Гетерополисахариды входят в состав межклеточного матрикса (основного вещества) соединительной ткани, участвуя в формировании протеогликанов.
В) Олигосахариды входят в состав гликопротеинов и гликолипидов – структурных компонентов плазматических и субклеточных мембран.
3. РЕГУЛЯТОРНАЯ.
А) Олигосахариды присутствуют в структуре многих гормонов, являющихся гликопротеинами.
Б) Белки-рецепторы для гормонов, расположенные на поверхности клеток, также являются гликопротеинами.
4. ЗАЩИТНАЯ. Примером может служить гетерополисахарид гепарин – ингибитор свертывания крови, препятствующий тромбообразованию.
МОНОСАХАРИДЫ
Содержат от трех до восьми атомов углерода (триозы, тетрозы, пентозы и т.д.). Среди моносахаридов можно выделить две группы: альдозы и кетозы. Альдозы (например, глюкоза), содержат альдегидную группу, а в составе кетоз (например, фруктоза) имеется кетогруппа. В природе встречаются L- и D- оптические изомеры, но в организме человека используются только D-стереоизомеры.
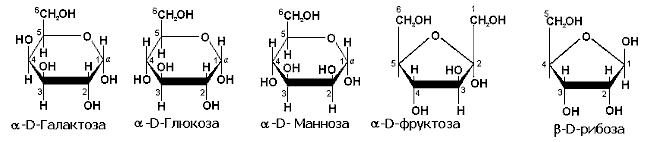
Рисунок XX. Строение некоторых главнейших моносахаридов организма человека.
ДИСАХАРИДЫ
Построены из двух моносахаридных фрагментов, соединенных гликозидной связью. Наиболее важное значение имеют сахароза (главный источник – пищевой сахар-рафинад) и лактоза (молочный сахар), поступающие в организм с пищей, а также мальтоза, образующаяся при переваривании пищевого крахмала или гликогена в желудочно-кишечном тракте.
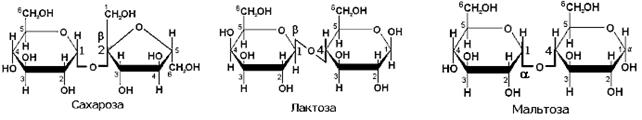
Рисунок ХХ. Строение важнейших дисахаридов
ОЛИГОСАХАРИДЫ
Содержат от трех до десяти моносахаридных фрагментов и, как правило, в свободном виде в организме человека не встречаются. Чаще всего они – компонент гликопротеинов и гликолипидов. Моносахаридные остатки так же, как и в дисахаридах, связаны друг с другом гликозидными связями.
ПОЛИСАХАРИДЫ
Молекулярная масса различных полисахаридов сильно варьирует, но в целом весьма высокая (от десятков тысяч до сотен миллионов дальтон). Связи между их многочисленными мономерными звеньями – такие же, как и у дисахаридов, и у олигосахаридов – гликозидные. Полисахариды могут принципиально отличаться друг от друга строением мономеров, и по этому признаку их можно разделить на две группы: гомополисахариды и гетерополисахариды. Мономером гомополисахаридов служит моносахарид. Наибольшее значение для организма человека представляют гомополисахариды крахмал, гликоген и, в меньшей степени, целлюлоза (клетчатка), мономером которых является глюкоза.
ГОМОПОЛИСАХАРИДЫ
Растительный сахар крахмал, являющийся для человека главным пищевым источником глюкозы – это смесь двух полисахаридов: амилозы, молекулы которой линейны, и амилопектина, обладающего разветвленной структурой.

Рисунок ХХ. Фрагмент молекулы амилозы.
Доля амилозы в крахмале составляет не более ¼, остальные ¾ приходятся на амилопектин.
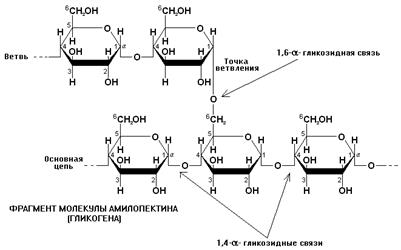
Рисунок ХХ. Фрагмент молекулы амилопектина или гликогена (животного крахмала).
Гликоген (животный крахмал) по структуре аналогичен амилопектину, но его молекулы обладают большей молекулярной массой и в два раза большей разветвленностью: ответвления от основной цепи встречаются в два раза чаще, чем в амилопектине. Поэтому по количеству остатков глюкозы, содержащихся в единице объема, гликоген в два раза превосходит амилопектин и обладает большей компактностью, что имеет важное значение из-за ограниченности внутреннего пространства в организме человека и животных. Гликоген синтезируется в биологически значимом количестве только в печени и в мышечной ткани, где выполняет функцию энергетического депо.
Еще одним гомополисахаридом, попадающим в организм человека в значительном количестве с растительной пищей, является целлюлоза (клетчатка). Неразветвленная молекула целлюлозы содержит примерно 10000 остатков β-глюкозы, соединенных друг с другом β(1,4)-гликозидными связями.
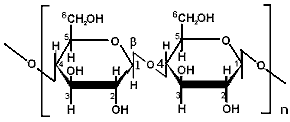
Рисунок ХХ. Фрагмент молекулы целлюлозы
В желудочно-кишечном тракте человека нет фермента, способного к расщеплению β-гликозидных связей. Однако целлюлоза, являясь неперевариваемым сахаром, тем не менее играет важную роль в пищеварении. Она стимулирует перистальтику желудочно-кишечного тракта, раздражая механорецепторы кишечной стенки, и способствует формированию каловых масс, сорбирует на себе токсические вещества, поступающие с пищей или образующиеся под действием ферментов микрофлоры кишечника.
ГЕТЕРОПОЛИСАХАРИДЫ
Мономером гетерополисахаридов является дисахарид, в котором моносахаридные фрагменты (как правило, это производные моносахаридов) связаны β(1,3)-гликозидной связью. Сами же мономеры образуют между собой β(1,4)-гликозидные связи. К полисахаридам этой группы относятся гиалуроновая кислота, хондроитинсульфаты, кератансульфат, дерматансульфат, гепаран-сульфат и гепарин. Мономеры многих из них содержат уроновые кислоты, почти все сульфатированы (кроме гиалуроновой кислоты).

Рисунок ХХ. Мономер хондроитин-4-сульфата, построенный из глюкуроновой кислоты и N-ацетилгалактозаминсульфата, соединенных β(1,3)-гликозидной связью.
Диссоциация карброксильной группы и остатка серной кислоты придает молекулам гетерополисахаридов значительный отрицательный заряд. Поэтому они сильно гидратированы и образуют гели. Принимая участие в образовании межклеточного матрикса соединительной ткани, гетерополисахариды обеспечивают его избирательную проницаемость для различных молекул (функция «молекулярного сита»). Кроме антикоагулянта гепарина, продуцируемого тучными клетками, остальные гетерополисахариды синтезируются фибробластами.
СИНТЕЗ И РАСПАД ГЛИКОГЕНА
Гликоген – полисахарид, накапливающийся, главным образом, в печени и в мышцах, и выполняющий функцию энергетического резерва углеводов, который может быть быстро использован при увеличении энергопотребления. Синтез гликогена протекает лишь в условиях «энергетического комфорта», когда количество поступающих углеводов с пищей превышает энергетические потребности клеток организма.
В случае синтеза гликогена после образования глюкозо-6-фосфата (гексокиназная реакция) происходит внутримолекулярный перенос остатка фосфорной кислоты из 6-го положения в 1-е. При этом образуется глюкозо-1-фосфат.
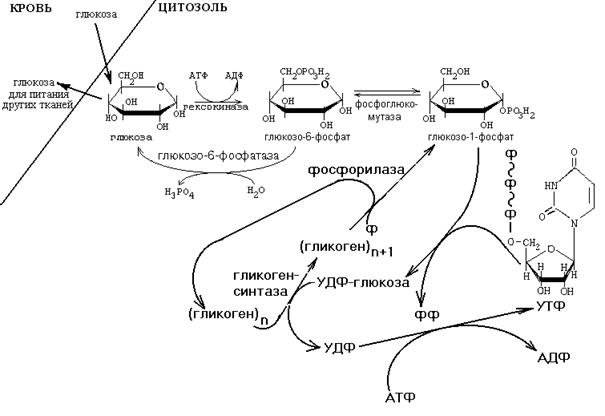
Рисунок ХХ. Схема метаболизма гликогена в печени
После изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат протекает дополнительная активация глюкозного фрагмента. При этом расходуется 1 молекула УТФ, что равнозначно затрате одной молекулы АТФ. В результате образуется УДФ-глюкоза.
Затем глюкозный остаток переносится на молекулу гликогена, и его цепь становится на один глюкозный фрагмент длиннее. Удлинение цепи гликогена катализирует фермент гликогенсинтетаза – ключевой фермент синтеза гликогена, который активируется избытком глюкозо-6-фосфата. Поэтому, если глюкозо-6-фосфат в клетке потребляется медленно, то возрастание его концентрации приводит к увеличению скорости синтеза гликогена. Реакция, катализируемая гликогенсинтазой, необратима.
Гликоген – разветвленная молекула. Для формирования ответвлений (α(1,6)-гликозидных связей) существует специальный фермент - "гликогенветвящий фермент".
Молекула гликогена синтезируется не «с нуля», а происходит постепенное удлинение уже имеющегося фрагмента цепи – так называемой "затравки". И при распаде гликогена никогда не происходит полного разрушения его молекул – обязательно остаются «затравки» - олигосахаридные фрагменты, которые в момент запуска его синтеза являются матрицей для удлинения полисахаридных цепей и формирования новых отвтевлений.
Для включения одного остатка глюкозы в молекулу гликогена клетка расходует 2 молекулы АТФ. При распаде гликогена освобождается только Фн (неорганический фосфат).
При увеличении энергопотребления гликоген способен распадаться. Для этого существует свой обходной обратный путь. Его ключевым ферментом является гликогенфосфорилаза, который угнетается избытком АТФ, активируется избытком АДФ и АМФ. Этот фермент расщепляет молекулу гликогена с участием неорганического фосфата (Фн) до глюкозо-1-фосфата и гликогена, укороченного на один глюкозный фрагмент: (С6Н10О5)n + H3PO4 ----> (C6H10O5)n-1 + глюкозо-1-фосфат
АЭРОБНЫЙ РАСПАД УГЛЕВОДОВ
Это главный путь распада углеводов до конечных продуктов, а для клеток некоторых тканей является единственно возможным. В среднем аэробному распаду подвергается 70-75% глюкозы, поступающей в клетку, и для многих тканей этот самый протяженный путь распада углеводов является главным источником получения энергии в клетке. Получение энергии в виде АТФ и является главным биологическим значением аэробного распада углеводов.
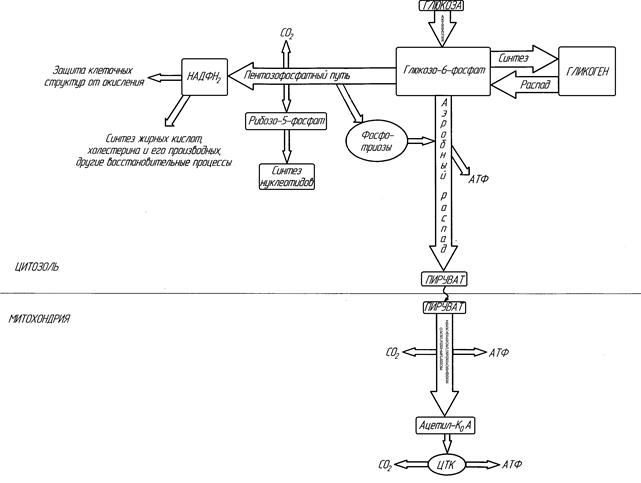
Рисунок ХХ. Главные пути метаболизма углеводов (схема).
Аэробный путь распада углеводов условно можно разделить на три этапа.
ГЛЮКОНЕОГЕНЕЗ
Некоторые промежуточные метаболиты аэробного пути распада углеводов могут образоваться из веществ других классов (не из углеводов): например, из аминокислот, липидов.
Приведем пример. ПВК может получиться в ходе реакции трансаминирования из аминокислоты аланина:
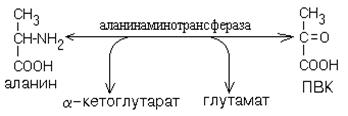
Рисунок ХХ. Образование пирувата из аланина.
После этого протекают все те же обратные реакции от пирувата до глюкозы (гликогена), как и при обращении гликолиза. Но поскольку пируват в данном случае имеет неуглеводное происхождение, то и процесс синтеза углеводов здесь называется по-другому: глюконеогенез.
ГЛЮКОНЕОГЕНЕЗ - это образование углеводов (например, глюкозы или гликогена) из веществ, имеющих неуглеводное происхождение.
Ключевым ферментом глюконеогенеза является пируваткарбоксилаза. В обычных условиях этот фермент малоактивен, но сильно активируется даже при небольшом накоплении АцетилКоА в цитоплазме. Тогда обходной обратный путь 10-й стадии и весь процесс синтеза углеводов из ПВК может протекать быстрее, чем их распад.
I. ОКИСЛИТЕЛЬНЫЙ ЭТАП
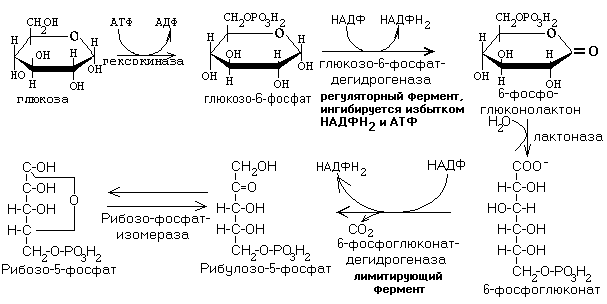
Рисунок ХХ. Реакции окислительного этапа пентозофосфатного пути распада углеводов.
На рисунке ХХ видно, что в случае пентозофосфатного пути после образования глюкозо-6-фосфата в гексокиназной реакции сразу же происходит его прямое окисление без участия кислорода. Сущность окислительного этапа заключается в окислении гексозофосфата (две реакции окисления, не требующие участия кислорода) и его декарбоксилировании, в результате чего образуется пентозофосфат. Отщепляемый при этом Н2 переносится на НАДФ с образованием НАДФН2. НАДФН2, образующийся в цитоплазме, не способен передавать свой водород в митохондрию на дыхательные цепи, и АТФ не образуется. Поэтому прямого энергетического значения пентозофосфатный путь для клетки не имеет. Водород НАДФН2 используется в восстановительных реакциях, протекающих при синтезе жирных кислот, холестерина и его производных, некоторых гормонов и биологически активных веществ для защиты клеточных структур от действия сильных окислителей и в некоторых других метаболические путях.
4. Образуются пентозы. Эти пентозы являются строительным материалом для синтеза нуклеотидов, коферментов и некоторых других веществ.
II. НЕОКИСЛИТЕЛЬНЫЙ ЭТАП.
Представляет собой совокупность большого количества обратимых реакций, каждая из которых сопровожлается переносом 2-х или 3-х углеродного фрагмента с одного моносахарида на другой. Вступают в него 6 молекул пентозо-фосфата одновременно. В результате взаимного обмена частями молекул образуются промежуточные продукты - моносахариды с разным числом углеродных атомов. Это – триозы (С3), тетрозы(С4), гексозы(С6), гептозы (С7).
Реакции неокислительного этапа катализируются ферментами трансальдолазами и транскетолазами. В состав кофермента транскетолаз входит витамин В1 (тиамин).
В ходе неокислительного этапа 6 молекул рибозо-5-фосфата превращаются в 5 молекул глюкозо-6-фосфата.
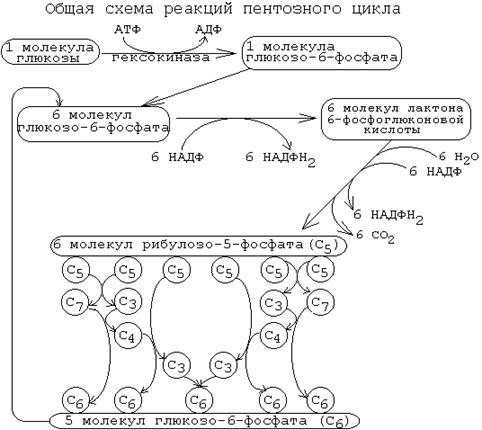
Таким образом, окислительный этап пентозофосфатного пути заключается в превращении 6 молекул глюкозы в 6 молекул глюкозо-6-фосфата, при двукратном окислении каждой из которых образуется рибозо-5-фосфат, 2 молекулы НАДФН2 и 1 молекула СО2. Затем, в ходе неокислительного этапа 6 молекул рибозо-5-фосфата трансформируются в 5 молекул глюкозо-6-фосфата. Эти 5 молекул глюкозо-6-фосфата вместе с еще одной, поступающей дополнительно, опять вступают в пентозофосфатный путь, замыкая цикл. На каждом "обороте" такого цикла происходит частичное окисление 6-ти молекул глюкозо-6-фосфата. В результате 5 молекул глюкозо-6-фосфата остаются нетронутыми, а одна молекула полностью расщепляется до СО2 и Н2О.
Итоговое уравнение пентозофосфатного пути:
С6Н12О6 + 6 Н2О + 12 НАДФ ------> 6 CO2 + 12 НАДФН2
Гормоны прямого действия.
ИНСУЛИН
Основные механизмы действия инсулина:
1. Инсулин повышает проницаемость плазматических мембран для глюкозы. Этот эффект инсулина является главным лимитирующим звеном метаболизма углеводов в клетках.
2. Инсулин снимает тормозящее действие глюкокортикостероидов на гексокиназу.
3. На генетическом уровне инсулин стимулирует биосинтез ферментов метаболизма углеводов, в том числе ключевых ферментов.
4. Инсулин в клетках жировой ткани ингибирует триглицеридлипазу - ключевой фермент распада жиров.
Регуляция секреции инсулина в кровь происходит с участием нейро-рефлекторных механизмов. В стенках кровеносных сосудов есть особые хеморецепторы, чувствительные к глюкозе. Повышение концентрации глюкозы в крови вызывает рефлекторную секрeцию инсулина в кровь, глюкоза проникает в клетки и ее концентрация в крови снижается.
Остальные гормоны вызывают повышение концентрации глюкозы в крови.
ГЛЮКАГОН.
Относится к белково-пептидным гормонам. Обладает мембранным типом взаимодействия с клеткой-мишенью. Эффект оказывает через аденилатциклазную систему.
1. Вызывает повышение активности гликоген-фосфорилазы. В результате ускоряется распад гликогена. Так как глюкагон оказывает эффект только в печени то можно сказать, что он "гонит глюкозу из печени".
2. Понижает активность гликоген-синтетазы, замедляя синтез гликогена.
3. Активирует липазу в жировых депо.
АДРЕНАЛИН.
Имеет рецепторы во многих тканях, а механизмы действия у него такие же, как у глюкагона.
1. Ускоряет распад гликогена.
2. Замедляет синтез гликогена.
3. Ускоряет липолиз.
ГЛЮКОКОРТИКОСТЕРОИДЫ (ГКС).
Относятся к стероидным гормонам, поэтому обладают внутриклеточным типом взаимодействия с клеткой-мишенью. Проникая в клетку-мишень, они взаимодействуют с клеточным рецептором и обладают следующими эффектами:
1. Ингибируют гексокиназу - таким образом они замедляют утилизацию глюкозы. В результате концентрация глюкозы в крови возрастает.
2. Данные гормоны обеспечивают процесс гликонеогенеза субстратами.
3. На генетическом уровне усиливают биосинтез ферментов катаболизма белков.
Гормоны косвенного действия
СОМАТОТРОПНЫЙ ГОРМОН.
1. Усиливает выделение глюкагона, поэтому наблюдается ускорение распада гликогена.
2. Вызывает активацию липолиза, поэтому способствует утилизации жира в качестве источника энергии.
ЙОДСОДЕРЖАЩИЕ ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ.
Трииодтиронин (Т3)
Тироксин (Т4)
Эти гормоны являются производными аминокислоты тирозина. Обладают внутриклеточным типом взаимодействия с клетками-мишенями. Рецептор Т3/Т4 находится в ядре клетки. Поэтому эти гормоны усиливают биосинтез белков на уровне транскрипции. Среди этих белков - окислительные ферменты, в частности разнообразные дегидрогеназы. Кроме того, они стимулируют синтез АТФаз - ферментов, которые разрушают АТФ. Для процессов биоокисления требуются субстраты - продукты окисления углеводов и жиров. Поэтому при увеличении продукции этих гормонов наблюдается усиление распада углеводов и жиров. Заболевание, при котором наблюдается гиперфункция щитовидной железы, называется Базедова болезнь (тиреотоксикоз). Один из симптомов этого заболевания - понижение массы тела. Для этого заболевания характерно повышение температуры тела. В опытах in vitro наблюдается разобщение митохондриального окисления и окислительного фосфорилирования при высоких дозах этих гормонов.
МЕТАБОЛИЗМ УГЛЕВОДОВ И ЕГО РЕГУЛЯЦИЯ
Углеводы – класс органических веществ, включающий в себя альдегиды и кетоны многоатомных спиртов, а также их производные. В природе углеводы существуют в форме моносахаридов, дисахаридов (в их составе – два моносахаридных фрагмента), олигосахаридов (содержат от трех до десяти моносахаридных фрагментов) и полисахаридов (содержат от десятков до десятков тысяч моносахаридных фрагментов).
Углеводы выполняют в организме несколько важных функций.




