ОБМЕН И ФУНКЦИИ УГЛЕВОДОВ
Функции углеводов
В основном углеводы выполняют энергодативную функцию. Главными источниками энергии являются глюкоза и гликоген. Кроме того, из углеводов могут синтезироваться липиды, некоторые аминокислоты, пентозы. Углеводы входят как составная часть в структурно-функциональные компоненты клетки - гликолипиды и гликопротеины, входят в состав соединительной ткани. Гепарин является антикоагулянтом.
Классификация углеводов
Моносахариды, которые не могут быть гидролизованы на более простые сахара. В зависимости от числа атомов углерода их подразделяют на триозы, тетрозы, пентозы, гексозы. В зависимости от присутствия альдегидной или кетоновой группы на альдозы и кетозы.
Дисахариды состоят из двух остатков моносахаридов:
1) сахароза состоит из остатков глюкозы и фруктозы, соединенных a-1,4-гликозидной связью;
2) лактоза состоит из остатков глюкозы и галактозы, соединенных b-1,4-гликозидной связью;
3) мальтоза состоит из двух остатков глюкозы, соединенных a-1,4-гликозидной связью;
4) целлобиоза состоит из двух остатков глюкозы, соединенных b-1,4-гликозидной связью.
Гомополисахариды - длинные разветвленные цепи, состоящие из одних и тех же моносахаридов:
1) крахмал - полимер глюкозы, соединенной a-1,4 и a-1,6-гликозидными связями. При этом неразветвленные цепи образуют амилозу (20%), а разветвленные амилопектин (80%);
2) гликоген - животный крахмал, состоящий из остатков глюкозы. Это более разветвленный полимер, чем крахмал. При частичном гидролизе крахмала или гликогена образуются декстрины (более короткие разветвленные цепи);
3) целлюлоза - главный компонент структурной основы растительных клеток. Это линейный полимер глюкозы, соединенной b-1,4-гликозидными связями.
Строение гликогена представлено на рисунке 1:

Рисунок 1
Гетерополисахариды состоят из разных мономеров:
1) гепарин содержит остатки D-глюконат-2-сульфита и N-ацетилглюкозамин-6-сульфата;
2) гиалуроновая кислота состоит из остатков D-глюкуроновой кислоты и остатков N-ацетилглюкозамина. Входит в состав соединительной ткани и участвует в регуляции проницаемости кканей.
Суточная норма углеводов в пище составляет 400-500 г.
Основными углеводами пищи являются:
1. крахмал - разветвленный гомополисахарид из глюкозы. Мономеры линейных участков соединены a -1,4-гликозидными связями, а в местах разветвления a -1,6 связями.
2. дисахариды - сахароза (глк-(a -1,2)-фру), лактоза (гал-(b -1,4)-глк), мальтоза (глк-(a -1,4)-глк).
Метаболизм глюкозы в клетке
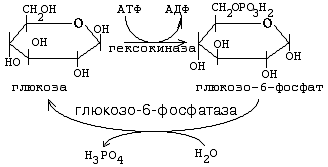
Первая реакция, в которую вступает глюкоза в клетке, является единственной. Это реакция фосфорилирования глюкозы за счёт АТФ. Фермент, катализирующий эту реакцию, есть в любой клетке. Он называется гексокиназа (ГК).
Биологический смысл гексокиназной реакции:
1. Сделать молекулу глюкозы более способной к химическим реакциям, ослабить в ней химические связи, дестабилизировать её ("расшатать").
2. Связать, задержать глюкозу в клетке, чтобы она не смогла выйти обратно в кровь (глюкозо-6-фосфат не способен проходить через клеточную мембрану).
Чтобы связанная молекула могла выйти из клетки, глюкозо-6-фосфат должен превратиться обратно в глюкозу. Фермент, катализирующий обратную реакцию (превращение глюкозо-6-фосфата обратно в глюкозу), называется глюкозо-6-фосфатаза. Он гидролизует глюкозо-6-фосфат до глюкозы и Н3РО4 (Фн), то есть катализирует обходной обратный путь гексокиназной реакции. Глюкозо-6-фосфатаза есть в печени, почках и слизистой оболочке кишечника.
3. Гексокиназа - это ключевой фермент всего метаболизма глюкозы. Он лимитирует (ограничивает) скорость всех путей метаболизма глюкозы в клетке. Гексокиназа всегда работает с максимальной скоростью.
Только в печени есть ещё один фермент, катализирующий реакцию превращения глюкозы в глюкозо-6-фосфат. Это изофермент гексокиназы - глюкокиназа. "На высоте пищеварения", когда концентрация глюкозы в воротной вене во много раз возрастает, глюкокиназа работает очень интенсивно. Следовательно, утилизация глюкозы клетками печени "на высоте пищеварения" возрастает при подключении дополнительного пути метаболизма (работа глюкокиназы).
Регуляторная роль гексокиназы: этот фермент угнетается избытком своего продукта - глюкозо-6-фосфата. Если по какой-то причине дальнейшее использование глюкозо-6-фосфата замедляется (его концентрация при этом возрастает), то автоматически тормозится гексокиназная реакция. Поэтому в такой ситуации замедляется использование в клетке глюкозы в целом.
После образования глюкозо-6-фосфата начинается разветвление дальнейших путей метаболизма глюкозы. Таких главных путей три.
1. Гексозомонофосфатный путь распада углеводов (ГМФ-путь)
2. Гексозобисфосфатный путь распада углеводов (ГБФ-путь).
3. Синтез гликогена.
Есть ещё минорные пути (в них используется небольшая доля глюкозы, поступающей в клетку). Эти пути не играют энергетической роли, а используются для построения олиго- и полисахаридных цепей гликопротеинов, то есть выполняют структурную роль.
СИНТЕЗ И РАСПАД ГЛИКОГЕНА.
Гликоген — основной резервный полисахарид в клетках животных и человека, так как он плохо растворим в воде и не влияет на осмотическое давление в клетке, поэтому в клетке депонируется гликоген, а не свободная глюкоза.
Разветвленная структура гликогена создает большое количество концевых мономеров. Это способствует работе ферментов, отщепляющих или присоединяющих мономеры при распаде или синтезе гликогена, так как эти ферменты могут одновременно работать на нескольких ветвях молекулы гликогена.
Гликоген депонируется главным образом в печени и скелетных мышцах. Гликоген хранится в цитозоле клеток в форме гранул. С гранулами связаны и некоторые ферменты, участвующие в обмене гликогена, что облегчает им взаимодействие с субстратом. Синтез и распад гликогена протекают разными метаболическими путями (рис 4).
Гликоген синтезируется в период пищеварения (1—2 ч после приема углеводной пищи). Синтез гликогена требует энергии. При включении одного мономера в
полисахаридную цепь протекают 2 реакции, сопряженные с расходованием АТР и UTP (реакции 1 и 3).
После образования глюкозо-6-фосфата (гексокиназная реакция) происходит внутримолекулярный перенос остатка фосфорной кислоты из 6-го положения в 1-е. При этом образуется глюкозо-1-фосфат
После изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат протекает дополнительная активация глюкозного фрагмента. При этом расходуется 1 молекула УТФ, что эквивалентно расходованию 1-й молекулы АТФ. В результате образуется активированная форма - УДФ-глюкоза (рис. 4).
Затем с УДФ глюкозный остаток переносится на молекулу гликогена. Удлинение цепи гликогена катализирует фермент гликогенсинтетаза. Таким образом, цепь гликогена становится на 1 глюкозный фрагмент длиннее. Гликоген, в отличие от растительного крахмала, более сильно разветвлен. Для формирования ответвлений существует специальный фермент, который называется "гликогенветвящий фермент".
Молекула гликогена синтезируется не с "нуля", а происходит постепенное удлинение уже имеющегося кусочка цепи: "затравки" или праймера. И при распаде гликогена никогда не происходит полного разрушения его молекул.
Для включения одного остатка глюкозы в молекулу гликогена клетка расходует 2 молекулы АТФ. При распаде гликогена эта АТФ не регенерирует, а освобождается только Фн (неорганический фосфат).
Ключевым ферментом синтеза гликогена является гликогенсинтаза. Это "пункт вторичного контроля" (рис. 5).
Регуляция гликогенсинтазы: она активируется избытком глюкозо-6-фосфата. Поэтому если глюкозо-6-фосфат другими путями утилизируется медленно, то возрастание его концентрации приводит к увеличению скорости синтеза гликогена. Реакция, катализируемая гликогенсинтазой, необратима.
Мобилизация гликогена происходит в основном в период между приемами пищи и ускоряется во врем физической работы. Этот процесс происходит путем последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата с помощью гликогенфосфорилазы (рис. 4). Этот фермент не расщепляет a1,6-гликозидные связи в местах разветвлений, поэтому необходимы еще 2 фермента, после действия которых глюкозный остаток в точке ветвления освобождается в форме свободной глюкозы (реакции 2, 3). Гликоген распадается до глюкозо-6-фосфата без затрат АТР.
Регуляция гликогенфосфорилазы: угнетается избытком АТФ, активируется избытком АДФ.
Распад гликогена в печени и мышцах имеет одну различающую их реакцию, обусловленную наличием в печени фермента фосфатазы глюкозо-6-фосфата (табл. 1).
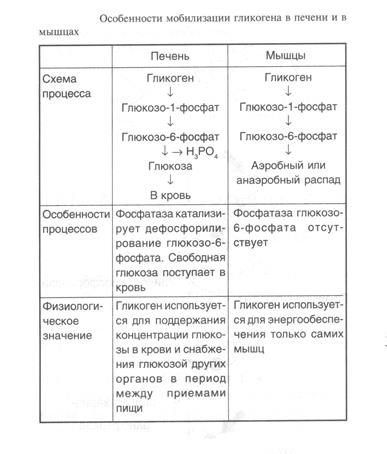
Таблица 1.
Присутствие в печени глюкозо-6-фосфатазы обусловливает главную функцию гликогена печени -освобождение глюкозы в кровь в период между приемами пищи и использование ее другими органами. Таким образом, мобилизация гликогена печени обеспечивает содержание глюкозы в крови на постоянном уровне. Это обстоятельство является обязательным условием для работы других органов и особенно мозга. Через 10—18 ч после приема пищи запасы гликогена в печени значительно истощаются, а голодание в течение 24 ч приводит к полному его исчезновению. Глюкозо-6-фосфатаза содержится также в почках и клетках кишечника.
Функция мышечного гликогена заключается в высвобождении глюкозо-6-фосфата, используемого в самой мышце для окисления и получения энергии,
Переключение процессов синтеза и мобилизации гликогена в печени происходит при переходе состояния пищеварения в постабсорбтивный период или состояния покоя на режим мышечной работы. В переключении этих метаболических путей в печени участвуют инсулин, глюкагон и адреналин, а в мышцах — инсулин и адреналин.
Влияние этих гормонов на синтез и распад гликогена осуществляется путем изменения в противоположном направлении активности 2 ключевых ферментов — гликогенсинтазы и гликогенфосфорилаз ы - с помощью их фосфорилирования и дефосфорилирования.
Первичным сигналом для синтеза инсулина и глюкагона является изменение концентрации глюкозы в крови. Инсулин и глюкагон постоянно присутствуют в крови, но при переходе из абсорбтивного состояния в постабсорбтивное изменяется их относительная концентрация — инсулин-глюкагоновый индекс. Таким образом, главным переключающим фактором в печени является инсулин-глюкагоновый индекс.
В постабсорбтивном периоде инсулин-глюкагоновый индекс снижается и решающим фактором является влияние глюкагона, который стимулирует распад гликогена в печени. Механизм действия глюкагона включает каскад реакций, приводящий к активации гликогенфосфорилазы.
В период пищеварения преобладающим является влияние инсулина, так как инсулин-глюкагоновый индекс в этом случае повышается. Под влиянием инсулина происходит:
а) стимуляция транспорта глюкозы в клетки мышечной ткани;
б) изменение активности ферментов путем фосфорилирования и дефосфорилирования. Так, например, инсулин активирует фосфодиэстеразу и снижает концентрацию сАМР в клетке. Кроме этого, инсулин активирует фосфатазу гликогенсинтазы, последняя дефосфорилируется и переходит в активное состояние;
в) изменение количества некоторых ферментов путем индукции и репрессии их синтеза. Например, инсулин индуцирует синтез глюкокиназы, ускоряя тем самым фосфорилирование глюкозы в печени.
Адреналин имеет сходный с глюкагоном механизм действия на клетки печени.Но возможно включение и другой эффекторной системы передачи сигнала в клетку печени. Тип рецепторов, с которыми взаимодействует адреналин, определяет, какая система будет использована. Так, взаимодействие адреналина с b-рецепторами клеток печени приводит в действие аденилатциклазную систему. Взаимодействие же адреналина с a,-рецепторами включает инозитолфосфатный механизм трансмембранной передачи гормонального сигнала. Результатом действия обеих систем являются фосфорилирование ключевых ферментов и переключение синтеза гликогена на его распад (рис.6, 7).
Активация адреналином мышечной гликогенфосфорилазы происходит иначе, так как распад гликогена в скелетных мышцах стимулируется мышечными сокращениями. Киназа фосфорилазы (Са2+-зависимая) активируется при мышечной работе под влиянием нервного импульса, так как в саркоплазме в этом случае возрастает концентрация ионов кальция. Это еще один механизм ускорения распада гликогена в мышце. Результатом действия адреналина в мышцах также являются активация сАМР-зависимых протеинкиназ и активация фосфорилазы путем ее фосфорилирования (рис. 8).
При передаче сигнала от гормона через внутриклеточные посредники происходит значительное его усиление, поэтому активация фосфорилазы гликогена при участии любой системы передачи сигнала в клетку позволяет быстро образовать большое количество глюкозы из гликогена. В мышцах это имеет большое значение для совершения интенсивной работы в условиях стресса, например при убегании от опасности.
При умеренной нагрузке в мышцах действует другой механизм регуляции активности гликогенфосфорилазы – аллостерическая регуляция продуктами распада АТФ (АМФ).
При переходе из постабсорбтивного состояния в абсорбтивное или по окончании мышечной работы прекращается секреция гормонов и вся система возвращается в исходное неактивное состояние. Аденилатциклаза и фосфолипаза С инактивируются. сАМР разрушается фосфодиэстеразой, что вызывает переход всех внутриклеточных ферментов каскада в неактивную форму.
Значение регуляции скоростей синтеза и распада гликогена в печени заключается в обеспечении постоянства концентрации глюкозы в крови. Регуляция обмена гликогена в мышцах обеспечивает энергетическим материалом как интенсивную работу мышц, так и энергозатраты в состоянии покоя.
Гормоны прямого действия.
ИНСУЛИН
Основные механизмы действия инсулина:
1. Инсулин повышает проницаемость плазматических мембран для глюкозы. Этот эффект инсулина является главным лимитирующим звеном метаболизма углеводов в клетках.
2. Инсулин снимает тормозящее действие глюкокортикостероидов на гексокиназу.
3. На генетическом уровне инсулин стимулирует биосинтез ферментов метаболизма углеводов, в том числе ключевых ферментов.
4. Инсулин в клетках жировой ткани ингибирует триглицеридлипазу - ключевой фермент распада жиров.
Регуляция секреции инсулина в кровь происходит с участием нейро-рефлекторных механизмов. В стенках кровеносных сосудов есть особые хеморецепторы, чувствительные к глюкозе. Повышение концентрации глюкозы в крови вызывает рефлекторную секркцию инсулина в кровь, глюкоза проникает в клетки и ее концентрация в крови снижается.
Остальные гормоны вызывают повышение концентрации глюкозы в крови.
ГЛЮКАГОН.
1. Вызывает повышение активности гликогенфосфорилазы. В результате ускоряется распад гликогена. Так как глюкагон оказывает эффект только в печени то можно сказать, что он "гонит глюкозу из печени".
2. Понижает активность гликогенсинтетазы, замедляя синтез гликогена.
3. Активирует липазу в жировых депо.
АДРЕНАЛИН.
Имеет рецепторы во многих тканях, а механизмы действия у него такие же, как у глюкагона.
1. Ускоряет распад гликогена.
2. Замедляет синтез гликогена.
3. Ускоряет липолиз.
ГЛЮКОКОРТИКОСТЕРОИДЫ (ГКС).
1. Ингибируют гексокиназу - таким образом они замедляют утилизацию глюкозы. В результате концентрация глюкозы в крови возрастает.
2. Данные гормоны обеспечивают процесс гликонеогенеза субстратами.
3. На генетическом уровне усиливают биосинтез ферментов катаболизма белков.
Гормоны косвенного действия
СОМАТОТРОПНЫЙ ГОРМОН.
1. Усиливает выделение глюкагона, поэтому наблюдается ускорение распада гликогена.
2. Вызывает активацию липолиза, поэтому способствует утилизации жира в качестве источника энергии.
ЙОДСОДЕРЖАЩИЕ ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ.
Поэтому эти гормоны усиливают биосинтез белков на уровне транскрипции. Среди этих белков - окислительные ферменты, в частности разнообразные дегидрогеназы. Кроме того, они стимулируют синтез АТФаз, т.е. ферментов, которые разрушают АТФ. Для процессов биоокисления требуются субстраты - продукты окисления углеводов и жиров. Поэтому при увеличении продукции этих гормонов наблюдается усиление распада углеводов и жиров.
ОБМЕН И ФУНКЦИИ УГЛЕВОДОВ
Функции углеводов
В основном углеводы выполняют энергодативную функцию. Главными источниками энергии являются глюкоза и гликоген. Кроме того, из углеводов могут синтезироваться липиды, некоторые аминокислоты, пентозы. Углеводы входят как составная часть в структурно-функциональные компоненты клетки - гликолипиды и гликопротеины, входят в состав соединительной ткани. Гепарин является антикоагулянтом.
Классификация углеводов
Моносахариды, которые не могут быть гидролизованы на более простые сахара. В зависимости от числа атомов углерода их подразделяют на триозы, тетрозы, пентозы, гексозы. В зависимости от присутствия альдегидной или кетоновой группы на альдозы и кетозы.
Дисахариды состоят из двух остатков моносахаридов:
1) сахароза состоит из остатков глюкозы и фруктозы, соединенных a-1,4-гликозидной связью;
2) лактоза состоит из остатков глюкозы и галактозы, соединенных b-1,4-гликозидной связью;
3) мальтоза состоит из двух остатков глюкозы, соединенных a-1,4-гликозидной связью;
4) целлобиоза состоит из двух остатков глюкозы, соединенных b-1,4-гликозидной связью.
Гомополисахариды - длинные разветвленные цепи, состоящие из одних и тех же моносахаридов:
1) крахмал - полимер глюкозы, соединенной a-1,4 и a-1,6-гликозидными связями. При этом неразветвленные цепи образуют амилозу (20%), а разветвленные амилопектин (80%);
2) гликоген - животный крахмал, состоящий из остатков глюкозы. Это более разветвленный полимер, чем крахмал. При частичном гидролизе крахмала или гликогена образуются декстрины (более короткие разветвленные цепи);
3) целлюлоза - главный компонент структурной основы растительных клеток. Это линейный полимер глюкозы, соединенной b-1,4-гликозидными связями.
Строение гликогена представлено на рисунке 1:
Рисунок 1
Гетерополисахариды состоят из разных мономеров:
1) гепарин содержит остатки D-глюконат-2-сульфита и N-ацетилглюкозамин-6-сульфата;
2) гиалуроновая кислота состоит из остатков D-глюкуроновой кислоты и остатков N-ацетилглюкозамина. Входит в состав соединительной ткани и участвует в регуляции проницаемости кканей.
Суточная норма углеводов в пище составляет 400-500 г.
Основными углеводами пищи являются:
1. крахмал - разветвленный гомополисахарид из глюкозы. Мономеры линейных участков соединены a -1,4-гликозидными связями, а в местах разветвления a -1,6 связями.
2. дисахариды - сахароза (глк-(a -1,2)-фру), лактоза (гал-(b -1,4)-глк), мальтоза (глк-(a -1,4)-глк).