Функциональная (изучает физико-химические процессы).
Выделяют разделы биохимии в зависимости от объекта изучения: биохимия животных, микроорганизмов, растений, человека, клиническая биохимия и т.д.
Основные задачи биохимии:
1. изучение процессов биокатализа;
2. изучение строения и функций нуклеиновых кислот;
3. изучение молекулярных механизмов наследственности;
4. изучение строения, обмена белков;
5. изучение превращения углеводов;
6. изучение процессов обмена липидов;
7. изучение роли биорегуляторов (гормоны, нейромедиаторы);
Изучение роли витаминов и минеральных веществ.
Значение БХ для медицины:
1. необходима для понимания сущности заболевания (патогенеза), его механизма. Пр.: сахарный диабет в результате недостатка инсулина, атеросклероз – нарушение обмена липопротеинов, опухолевый рост – функционирование онкогенов;
2. необходима для диагностики заболеваний. Пр.: биохимический анализ крови, мочи. Определяется:
а) количество субстрата (уровень метаболита);
б) активность фермента;
в) количество биорегуляторов (гормонов и нейропептидов);
В диагностике заболеваний используются различные методы: радио-иммуный анализ, иммуно-ферментный анализ, введение ДНК-зондов для выявления чужеродной ДНК, дефектов ДНК, онкогенов. Также позволяет выявить предрасположенность к заболеваниям;
3. разработка новых лекарственных препаратов;
Необходима для профилактики заболеваний. Пр.: рахит – в результате недостатка витамина D, цинга – витамина С.
Белки и их биологическая роль
Белок (протеины) – protos – предшествующий всему, первичный, наиглавнейший, определяющий всё остальное.
Белки – это высокомолекулярные азотсодержащие органические вещества, состоящие из аминокислот, соединённых в цепи с помощью пептидных связей и имеющих сложную структурную организацию.
Основные отличительные признаки белков:
1. содержат азота больше, чем другие вещества (16%). Так, 1г азота содержится в 6,25г белка;
2. состоят из альфа-аминокислот L-ряда;
3. наличие пептидных связей;
4. большая молекулярная масса (от 4-5 тыс. дальтон до нескольких млн.);
5. имеют сложную структурную организацию;
6. белки составляют 25% сырой ткани и 45-50% сухой ткани.
Биологическая роль белков:
1. каталитическая (выполняют ферменты);
2. структурная, т.е. белки являются основным компонентом клеточных структур;
3. регуляторная (выполняют белки-гормоны);
4. рецепторная, т.е. рецепторы клеточных мембран имеют белковую природу;
5. транспортная – белки участвуют в транспорте липидов, токсических веществ, кислорода и т.д.;
6. опорная – выполняет белок коллаген;
7. энергетическая. Заключается в том, что при окислении 1г белка выделяется 17,6 кДж (4,1ккал) энергии;
8. сократительная – её выполняют белки актин и миозин;
9. генно-регуляторная – её выполняют белки гистоны, участвуя в регуляции репликации;
10. имуннологическая – её выполняют белки антитела;
11. гемостатическая – участвуют в свёртывании крови, препятствуют кровотечению;
12. антитоксическая, т.е. белки связывают многие токсические вещества (особенно соли тяжёлых металлов) и препятствуют развитию интоксикации в организме.
Физико-химические свойства белков:
Наличие заряда используется для разделения белков с помощью электрофореза – движения белков в электрическом поле. Наличие заряда обусловливает устойчивость в растворе. В изоэлектрическом состоянии белки наименее устойчивы и выпадают в осадок.
II. Коллоидные свойства.
Растворы белков чаще всего достаточно устойчивы. Хорошая растворимость приближает растворы белков к истинным растворам, но высокая молекулярная масса придает им свойства коллоидных систем:
1. способность рассеивать свет (опалисценция). Наблюдается помутнение при боковом освещении - эффект Тиндаля [рис. рассеивающегося луча]. Используется в световой микроскопии (нефелометрии);
2. малая скорость диффузии;
3. высокая вязкость растворов белков;
4. неспособность белков проникать через полупроницаемые мембраны (явление осмоса). На этом основан диализ – очищение белков;
Под действием внешних факторов нарушается высшие уровни (вторичный, третичный, четвертичный) структурной организации белков с сохранением первичной структуры. При этом белок теряет свои нативные свойства. При денатурации разрываются связи, удерживающие высшие структурные организации. Денатурацию вызывают физические и химические факторы: давление, температура, механическое воздействие, ультразвук, ионизирующее излучение, кислоты, щёлочи, органические растворители, соли тяжёлых металлов. При кратковременном воздействии денатурирующих факторов возможна ренатурация.
Классификация белков
В настоящее время насчитывается ~5 млн. белков. Их пытались классифицировать по физико-химическим свойствам, например по растворимости, плотности, форме молекул (глобулярные и фибриллярные), локализации и происхождению, АК-составу, биологической роли. Однако все эти классификации не соответствуют тем знаниям о белках, которые известны на сегодняшний день.
В основе классификации лежит химический состав белка. По этому признаку все белки делят на простые и сложные.
Простые белки – это белки, образованные только полипептидными цепями, состоящие только из АК-ных остатков.
Сложные белки имеют две части: белковая или пептидная построена из АК-ных остатков, и небелковая (простетическая) часть.
Они имеют белковую и небелковую (простетическую) части. Белковую часть составляет полипептид, построенный из АК-остатков. В состав небелковой части может входить: гем, металл, остаток фосфорной кислоты, углеводы, липиды и т.д.
Хромопротеины
Для них простетическая часть окрашена (chromos – краска). К хромопротеинам относятся гемоглобин, миоглобин, каталаза, пероксидаза, ряд флавинсодержащих ферментов (сукцинатдегидрогеназа, альдегидоксидаза, ксантиноксидаза), цитохромы (гемсодержащие белки) и т.д. Велика биологическая роль этих белков – участвуют в физиологических процессах: дыхание клетки, транспорте кислорода и углекислого газа, окислительно-восстановительных процессах.
Липид-белковые комплексы
Липид-белковые комплексы – сложные белки, простетическую часть которых составляют различные липидные компоненты. К таким компонентам относятся:
1. предельные и непредельные ВЖК. К предельным относятся стеариновая С17Н35СООН и пальмитиновая С15Н31СООН; к непредельным – олеиновая С17Н33СООН, линолевая С17Н31СООН, линоленовая С17Н29СООН и др.;
2. простые и смешанные триацилглицериды. К простым относится трипальмитин, а к смешанным – олеодистеарин:
СН2 - О - СО - С15Н31 СН2 - О - СО - С17Н33
| |
СН - О - СО - С15Н31 СН - О - СО - С17Н35
| |
СН2 - О - СО - С15Н31 СН2 - О - СО - С17Н35
Биологическая роль свободных ЛП сводится к транспортным функциям. За счет их гидрофильной оболочки происходит перенос различных веществ в клетки. Также они играют важную роль в диагностике патогенеза.
Нуклеопротеины
Нуклеопротеины – это сложные белки, содержащие в качестве небольшой части нуклеиновые кислоты (до 65%).
НП состоят из 2-х частей: белковой (содержит гистоны и протамины, которые являясь основными белками, придают основные свойства) и простетической, представленной НК, сообщающими кислотные свойства. Взаимодействие между этими частями по ион-ионному механизму.
НК – высокомолекулярные органические вещества, полинуклеотиды. Мономерами являются мононуклеотиды. Каждый мононуклеотид состоит из: углевода, азотистого основания и фосфорной кислоты. Так, РНК содержит b -D-рибофуранозу (рибозу), одно из 4-х возможных азотистых оснований (А, Г, Ц или У) и остаток фосфорной кислоты. ДНК содержит b -D-дезоксирибофуранозу (дезоксирибозу), одно из 4-х возможных азотистых оснований (А, Г, Ц или Т) и остаток фосфорной кислоты.
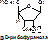
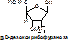
Строение азотистых оснований:
Молекула спирализована на всем протяжении, гидрофобные участки внутри спирали, их плоскости перпендикулярны основаниям и параллельны друг другу. В вертикальном направлении возникают гидрофобные взаимодействия. Вторичная структура стабилизируется водородными связями и гидрофобными взаимодействиями.
Вторичная структура РНК более простая, представляет собой одну полинуклеотидную цепь, в которой спирализованы лишь некоторые участки. Вторичная структура РНК представлена в виде клеверного листа. Для тРНК известна третичная структура в форме буквы Г. [рис. РНК в виде клеверного листа]
Биологическая роль НК:
ДНК – основная часть её локализуется в ядре в виде ДНП в составе хроматина или хромосом делящихся клеток. Главная роль – хранение генетической информации, участие в процессе транскрипции в качестве матрицы для построения молекулы РНК.
Все РНК по функции делятся на:
- рРНК (рибосомальные), составляют до 80% в составе рибосом. Играют роль каркаса для объединения рибосом белков;
- мРНК (иРНК) – образуется в ядре (ядрышке). Переносит информацию из ядра в цитоплазму, является матрицей в процессе трансляции белка. последний кодон иРНК соответствует последней АК в белке;
- тРНК по своей форме напоминает форму клеверного листа и представляет собой полинуклеотидную цепь, которая составляет 3 петли и отдельные участки могут быть спирализованы. тРНК активирует аминокислоты и транспортирует их к месту биосинтеза белков, также участвует в трансляции. Имеет антикодоновый триплет – место, с помощью которого тРНК связывается с комплиментарным кодоном мРНК.
Углевод-белковые комплексы
Гепарин не является структурным компонентом межклеточного матрикса. Он вырабатывается тучными клетками соединительной ткани и после их цитолиза выделяется в межклеточное пространство и кровеносное русло. В крови может соединяться с неспецифическими белками. Гепарин – естественный природный коагулянт, препятствует свёртыванию крови.
ПГ выполняют следующую роль:
1. роль «рессор» – смягчают нагрузки на суставные поверхности (в хрящах, суставных поверхностях);
2. ограничивают диффузию и проницаемость межклеточного вещества для различных молекул и крупных частиц. Гиалуроновая кислота подвергается гидролизу под действием фермента – гиалуронидазы. В норме, в здоровой ткани, фермент и субстрат находятся в равновесии. при патологических процессах количество фермента может увеличится, что приводит к гидролизу гиалуроновой кислоты и нарушению проницаемости. При этом разрываются b -1,4-гликозидные связи;
3. являются поливалентными анионами. Способны связывать большие количества ионов Na+ и др. катионов и участвуют в регуляции водно-солевого обмена;
4. Гиалуроновая кислота может присутствовать в свободном виде, например в хрусталике глаза, суставной жидкости. Она имеет консистенцию геля и выполняет различные функции.
Фосфопротеины
Белки, где в качестве простетической группы – фосфорная кислота. Присоединение фосфорной кислоты к полипептидной цепи идет с образованием сложноэфирной связи с АК СЕР или ТРЕ.
Типичными фосфопротеинами являются: казеин молока (1% фосфорной кислоты), желток куриного яйца (вителлин), икра рыб (ихтулин). Большое количество ФП содержится в нервных клетках. Для ФП характерен широкий диапазон функций в метаболизме. За счет фосфорилирования и дефосфорилирования ферментов происходит регуляция их активности.
ФЕРМЕНТЫ
Ферменты – это биологические катализаторы белковой природы. В одной клетке до 10 тыс. молекул фермента, которые катализируют 2000 ферментативных реакций. 1800 тыс. ферментов выделены, но их строение не расшифровано. Старое название ферментов – энзимы, а наука, их изучающая – энзимология.
По своей химической природе ферменты – это белки, они имеют несколько уровней структурной организации и обладают всеми другими свойствами белков. Очень большое количество ферментов не имеют 4-ую структуру, т.е. являются олигомерами.
Ферменты могут быть простыми и сложными. Простые состоят только из полипептидной цепи, а сложные имеют пептидную (апофермент) и небелковую части (кофермент). Далее идёт рис. [апофермент+кофермент=холофермент – обладает полноценной функциональной активностью]. По отдельности ни апофермент, ни кофермент, не выполняют функции так, как холофермент.
Строение ферментов. В пространственной структуре фермента условно выделяют ряд участков, которые выполняют соответствующие им функции. Активный центр (АЦ) – участок в молекуле фермента, где происходит связывание и химическое превращение субстрата (S). Субстрат – вещество, подвергающееся химическому превращению (например, для фермента лактатдегидрогеназы (ЛДГ) субстратом будет молочная кислота). В активном центре выделяется контактный участок и каталитический участок. Контактный участок – это место активного центра, в котором происходит связывание фермента с субстратом по принципу комплементарности, т.е. именно контактный участок обеспечивает специфическое сродство субстрата ферменту. Образовавшийся комплекс носит название фермент-субстратный комплекс. Каталитический участок (центр) – это место в активном центре фермента, где происходит химическое превращение субстрата [рис. изображён фермент-субстратный комплекс, а именно контактный участок, каталитический участок, активный центр и субстрат].
Если фермент – сложный белок, то обычно простетическая часть находится тоже в активном центре и участвует в формировании активного центра. Активный центр занимает небольшую часть молекулы фермента, обычно располагается в углублении, и в его образовании участвует небольшое число аминокислотных остатков (до 20). Аминокислотные остатки могут быть удалены друг от друга, но при формировании пространственной структуры фермента они располагаются в области активного центра.
В формировании активного центра могут участвовать остатки, несущие следующие функциональные группы: NH2 (ЛИЗ, АРГ), COOH (ГЛУ, АСП), OH (СЕР, ТРЕ), SH (ЦИС), имидазольное кольцо (ГИС). В качестве единиц, участвующих в формировании активного центра, могут выступать кофакторы - ионы металлов (Cu2+, Fe2+ и т.д.), а также коферменты. В сложном ферменте АК-остатки активного центра создают условия для правильной его конформации и помогают кофакторам в связывании, ориентации, а, следовательно, и в превращении субстрата.
Боковые группы остальных аминокислот не участвуют в образовании активного центра, но обеспечивают правильную пространственную конформацию активного центра и влияют на его реакционную способность.
Ряд ферментов могут содержать аллостерический центр. [рис. фермента с аллостерическим и активным центрами] Эти ферменты относят к аллостерическим ферментам. К аллостерическому центру присоединяются различные вещества, отличные по строению от субстрата. Эти вещества могут изменять конформацию активного центра, т.е. влиять на связывание и превращение субстрата, они называются аллостерическими эффекторами. Все аллостерические эффекторы делятся на положительные – активаторы, и отрицательные – ингибиторы.
Строение коферментов
Коферменты в каталитических реакциях осуществляют транспорт различных групп атомов, электронов или протонов. Коферменты связываются с ферментами:
- ковалентными связями;
- ионными связями;
- гидрофобными взаимодействиями и т.д.
Один кофермент может быть коферментом для нескольких ферментов. Многие коферменты являются полифункциональными (например, НАД, ПФ). В зависимости от апофермента зависит специфичность холофермента.
Изоферменты
Изоферменты – это изофункциональные белки. Они катализируют одну и ту же реакцию, но отличаются по некоторым функциональным свойствам в силу отличий по:
- аминокислотному составу;
- электрофоретической подвижности;
- молекулярной массе;
- кинетике ферментативных реакций;
- способу регуляции;
- стабильности и др.
Примеры изоферментов: глюкокиназа и гексокиназа.
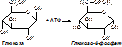 +АДФ
+АДФ
Гексокиназа может фосфорилировать любой шестичленный цикл, гексокиназа – только превращение глюкозы. После приёма пищи, богатой глюкозой, глюкокиназа начинает работать. Гексокиназа – стационарный фермент. Он катализирует реакцию расщепления глюкозы при низких её концентрациях, поступающих в организм. Отличаются по локализации (глюкокиназа – в печени, гексокиназа – в мышцах и печени), физиологическому значению, константе Михаэльса.
Если фермент – олигомерный белок, то изоформы могут получаться в результате различной комбинации протомеров. Например, лактатдегидрогеназа состоит из 4-х субъединиц. Н – субъединицы сердечного типа, М – мышечного. Может быть 5 комбинаций этих субъединиц, а, следовательно, и 5 изоферментов: НННН (ЛДГ1 – в сердечной мышце), НННМ (ЛДГ2), ННММ (ЛДГ3), НМММ (ЛДГ4), ММММ (ЛДГ5 – в печени и мышцах). [рис. эти 4 буквы в кружочки.
Надо отличать изоферменты от множественных форм ферментов. Множественные формы ферментов – это ферменты, которые модифицированы после своего синтеза, например фосфорилаза A и B.
Свойства ферментов
Общие черты ферментов и небиологических катализаторов:
1) и те, и другие катализируют только энергетически возможные реакции;
2) увеличивают скорость реакции;
3) не меняют направления реакции;
4) в ходе реакции не расходуются;
Повышение активности фермента при увеличении температуры объясняется увеличением кинетической энергии реагирующих молекул, что приводит к увеличению числа столкновений между молекулами. При дальнейшем повышении температуры энергия становится чрезмерной, и внутри молекулы разрываются слабые связи – водородные, гидрофильные взаимодействия; происходит нарушение вторичной, третичной, четвертичной структуры фермента.
Ряд ферментов термостабильны, например, гликопротеины.
4) Влияние рН. Для поддержания третичной или четвертичной структуры фермента часто может быть необходимо наличие заряда на группе, удаленной от области связывания субстрата. Если же заряд этой группы меняется, то может происходить частичное развертывание белковой цепи, или компактизация, или диссоциация (олигомерные белки). Поэтому при отклонении рН от оптимального значения, фермент может потерять свою нативную структуру, в результате чего не происходит полноценного связывания активного центра с субстратом. Также при изменении рН может происходить изменение заряда на субстрате.
[рис. график. По х – рН, по у – процент активности. Рисуем горочку.]
Лигазы (синтетазы).
Пр.: каталазная реакция
2Н2О2 → 2Н2О+О2 [над стрелочкой - каталаза].
2. Трансферазы – ферменты, переносящие группы атомов от одного субстрата к другому. При этом один субстрат донор, а другой – акцептор. В зависимости от природы переносимых групп трансферазы делят на 8 подклассов:
- аминотрансферазы, переносят NH2;
- метилтрансферазы, переносят CH3;
- фосфотрансферазы, переносят PO3H2;
- ацилтрнсферазы.
Пр. переаминирование. [рис. Аланин + альфакетоглутаровая кислота под действием АлАТ и перидоксальфосфата получается ПВК и глутаминовая к-та].
3. Гидролазы – ферменты, катализирующие разрыв одинарных связей с участием воды, присоединяемой по месту разрыва связи. Т.е. они принимают участие в реакциях гидролиза. Все ферменты пищеварительного тракта относятся к гидролазам. Всего выделяют 460 гидролаз. В зависимости от типа разрываемых связей выделяют 11 подклассов:
- эстеразы – разрывают сложно-эфирную связь. Пр.: триацилглицерид+ 3H2O=(липаза) глицерин+ 3С17H35COOH [рис. этого. Стрелочкой показать сложноэфирные связи]
- пептидазы – разрывают пептидную связь. Пр.: АЛА-ГЛИ+ H2O = (дипептидаза) АЛА+ГЛИ [рис. этого. Стрелочкой показать пептидную связь]
- гликозидазы – разрывают гликозидные связи. Пр.: мальтоза+ H2O=(мальтаза) 2глюкозы [рис. этого. Стрелочкой показать гликозидную связь]
4) Лиазы. Эти ферменты осуществляют разрыв углеродных связей без участия воды. Выделяют:
- декарбоксилазы – катализируют отщепление CO2. Пр.:
- альдолазы – катализируют расщепление связи между атомами углерода. Пр.: фруктозо-1,6-дифосфат (6 атомов С)= (альдолаза) фосфоглицериновый альдегид (3 атома С)+ дигидроксиацетонфосфат (3 атома С).
- гидратазы – разрыв двойной связи с присоединением воды по месту разрыва двойной связи.
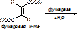
- ферменты, отщепляющие воду – реакция дегидратации. При этом образуется двойная связь.
Номенклатура ферментов
1) Существует тривиальная номенклатура – названия случайные, без системы и основания, например трипсин, пепсин, химотрипсин.
2) Рабочая номенклатура – название фермента составляется из названия субстрата или продукта реакции, типа катализируемой реакции и окончание –аза, например лактатдегирогеназа.
Первая теория ферментативного катализа была выдвинута в начале 20 века Варбургом и Бейлисом. Эта теория предлагала считать, что фермент адсорбирует на себе субстрат, и называлась адсорбционной, но развития она не получила.
В основу современной теории положена теория Михаэлеса и Ментена. Ведущую роль в механизме ферментативного катализа играет образование фермент-субстратного комплекса. По этой теории весь процесс катализа можно разделить на 3 этапа:
[ рис. E+S «ES ® ES* ® ES** ® EP ® E+P]
1 этап: образование фермент-субстратного комплекса (на рис. до ES включительно). Происходит диффузия субстрата к ферменту и субстрат, в соответствии с принципом комплиментарности, связывается с активным центром фермента – образуется фермент-субстратный комплекс. Реагенты связаны слабыми связями, т.е. водородными, ионными, гидрофобными, в некоторых случаях и ковалентными. Эта стадия непродолжительна, зависит от концентрации субстрата и от скорости диффузии его к активному центру. Энергия активации исходных веществ при этом изменяется незначительно. На этой стадии проявляется эффект концентрирования субстрата на поверхности фермента – эффект ориентации.
2 этап: (на рис. от ES до EP включительно) происходит последовательное преобразование первичного фермент-субстратного комплекса в 1 или несколько активированных. Эта стадия наиболее медленна, ее длительность зависит от величины энергии активации данной реакции. В эту стадию происходит разрыв старых связей и образование новых, при этом энергия активации значительно снижается. По продолжительности эта стадия является лимитирующей для всего процесса.
На этой стадии проявляется эффект вынужденного соответствия – эффект «дыбы»: субстрат под действием фермента претерпевает изменения, делающие его более доступным для воздействия каталитического участка активного центра фермента. Одновременно с этим происходит изменение конформации фермента в большей степени в активном центре.
Эффект вынужденного соответствия (прилегания). Не только субстрат претерпевает изменение конформации, но и фермент, особенно в активном центре, после связывания субстрата меняет свою конформацию, которая становится более комплементарной субстрату.
Теория Котланда: фермент и субстрат взаимодействуют между собой по принципу рука–перчатка. Истинная комплементарность фермента к субстрату достигается после изменения конформации и субстрата и фермента.
В составе активного центра фермента имеются как кислые, так и основные функциональные группы. В результате этого фермент проявляет в ходе катализа кислотно-основные свойства, т.е. играет как роль донора, так и роль акцептора протонов. Кислотно-основной катализ характерен для гидролаз, лиаз, изомераз.
При закреплении субстрата в активном центре на его молекулу влияют электрофильные и нуклеофильные группы каталитического участка, что вызывает перераспределение электронной плотности в субстрате. Такое перераспределение облегчает перестройку и разрыв связей в молекуле субстрата.
Пр.: Реакция превращения ацетилхолина в холин. На первом этапе между СОО- глутамина и N + ацетилхолина возникает ионная связь, и возникает фермент-субстратный комплекс. Начинается вторая стадия.
После образования фермент-субстратного комплекса в действие вступают остальные аминокислоты, остатки активного центра. Между углеродом С=О группы ацетилхолина и кислородом ОН-группы серина возникает взаимодействие, т.е. возникает водородная связь между кислородом ацетилхолина и ОН-группы тирозина – эффект «дыбы».
Затем гистидин оттягивает протоны от ОН-группы серина. Вследствие этого упрочняется сложноэфирная связь между серином и остатком уксусной кислоты. Одновременно происходит разрыв другой сложноэфирной связи в молекуле ацетилхолина и переход протона от тирозина к остатку холина.
На третьем этапе холин высвобождается из активного центра. Его место занимает вода. Эта вода располагается между карбонильным кислородом ацетильной группы и кислородом тирозина. Фермент освобождён от продуктов реакции и готов к следующему циклу. На первом и последнем этапе продолжительность этапа зависит от скорости диффузии субстрата к ферменту или от фермента соответственно. Вторая стадия очень часто является лимитирующей весь процесс. Именно на этой стадии происходит снижение энергии активации реагирующих веществ.
Активация ферментов. Различные активаторы могут связываться либо с активным центром фермента, либо вне его. К группе активаторов, влияющих на активный центр, относятся: ионы металла, коферменты, сами субстраты.
Активация с помощью металлов протекает по различным механизмам:
- металл входит в состав каталитического участка активного центра;
- металл с субстратом образуют комплекс;
- за счет металла образуется мости между субстратом и активным центром фермента.
Частичный протеолиз (ограниченный протеолиз). Таким образом активируются ферменты пищеварительного канала: пепсин, трипсин, химотрипсин. Трипсин имеет состояние профермента трипсиногена, состоящего из 229 АК остатков. Под действием фермента энтерокиназы и с добавлением воды он превращается в трипсин, при этом отщепляется гексапептид. Изменяется третичная структура белка, формируется активный центр фермента и он переходит в активную форму.
2) фосфорилирование - дефосфорилирование. Пр.: липаза+АТФ= (протеинкиназа) фосфорилированная липаза+АДФ. Это трансферная реакция, использующая фосфат АТФ. При этом осуществляется перенос группы атомов от одной молекулы к другой. Фосфорилированная липаза является активной формой фермента.
Таким же путем происходит активация фосфорилазы: фосфорилаза B+ 4АТФ= фосфорилаза А+ 4АДФ
Также при связывании активатора вне активного центра происходит диссоциация неактивного комплекса «белок-активный фермент». Например, протеинкиназа – фермент, осуществляющий фосфорилирование (цАМФ-зависимое). Протеинкиназа – это белок, имеющий четвертичную структуру и состоящий из 2-х регуляторный и 2-х каталитических субъединиц. R2C2+2цАМФ=R2цАМФ2+ 2С. Такой тип регуляции называется аллостерической регуляцией (активацией).
Ингибирование ферментов. Ингибитор – это вещество, вызывающее специфическое снижение активности фермента. Следует различать ингибирование и инактивацию. Инактивация – это, например, денатурация белка в результате действия денатурирующих агентов.
Аллостерическое ингибирование – торможение ферментативной реакции, вызванное присоединением аллостерического ингибитора в аллостерическом центре аллостерического фермента. Такой тип ингибирования характерен для аллостерических ферментов, имеющих четвертичную структуру. В качестве ингибиторов могут выступать метаболиты, гормоны, ионы металлов, коферменты.
Механизм действия:
а) присоединение ингибитора к аллостерическому центру;
б) изменяется конформация фермента;
в) изменяется конформация активного центра;
г) нарушается комплиментарность активного центра фермента к субстрату;
д) уменьшается число молекул ES;
Обмен веществ
Обмен веществ (метаболизм) – это совокупность физиологических и биохимических процессов, обеспечивающих жизнедеятельность организма во взаимосвязях с внешней средой, направленных на самовоспроизведение и самосохранение.
К физиологическим процессам относятся пищеварение, всасывание, внешнее дыхание, выделение и др.; к биохимическим – химические превращения белков, жиров, углеводов, поступающих в организм в виде пищевых веществ. Особенностью биохимических процессов является то, что они осуществляются в ходе ряда ферментативных реакций. Именно ферменты обеспечивают определенную последовательность, места и скорость реакций.
По направленности все химические превращения делят на:
а) диссимиляция (катаболизм) – распад веществ до более простых с переходом энергии связей вещества в энергию макроэргических связей (АТФ, НАД·Н, др.);
б) ассимиляция (анаболизм) – синтез более сложных веществ из более простых с затратой энергии.
Биологическое значение этих двух процессов состоит в том, что при расщеплении веществ освобождается заключенная в них энергия, которая обеспечивает все функциональные возможности организма. В то же время, при распаде веществ образуются "строительные материалы" (моносахариды, АК, глицерин и др.), которые затем используются в синтезе специфических организму веществ (белков, жиров, углеводов и др.).
[СХЕМА] Над горизонтальной линией (во внешней среде) – "белки, жиры, углеводы", от них стрелка вниз под линию (внутри организма) к надписи "диссимиляция", от последней четыре стрелки: две вверх к надписям над линией "теплота" и "конечные продукты"; одна стрелка вправо к надписи "промежуточные вещества (метаболиты)", от них к "ассимиляция", затем к "собственные белки, жиры, углеводы"; одна стрелка вниз к надписи "энергия АТФ", от нее – к "мышечное сокращение, проведение нервного импульса, секреция и др." а также наверх к "теплота" и "ассимиляция".
Диссимиляция белков, жиров и углеводов протекает по-разному, но в разрушении этих веществ есть ряд общих этапов:
1) Этап переваривания. В ЖКТ белки распадаются до АК, жиры – до глицерина и ВЖК, углеводы – до моносахаридов. Нарабатывается большое количество неспецифических веществ из специфических, поступающих извне. За счет переваривания в ЖКТ выделяется около 1% химической энергии веществ. Этот этап необходим для того, чтобы вещества, поступившие с пищей, смогли всосаться.
2) Этап межуточного обмена (тканевой обмен веществ, метаболизм). На клеточном уровне он распределяется на анаболизм и катаболизм. Образуются и превращаются промежуточные вещества обмена веществ – метаболиты. При этом мономеры, образовавшиеся на этапе переваривания, распадаются с образованием небольшого (до пяти) ключевых промежуточных продуктов: ЩУК, альфа-КГ, ацетил-КоА, ПВК, альфа-глицерофосфат. Выделяется до 20% энергии веществ. Как правило, межуточный обмен происходит в цитоплазме клеток.
3) Окончательный распад веществ с участием кислорода до конечных продуктов (СО2, Н2О, азотсодержащие вещества). Выделяется около 80% энергии веществ.
Все рассмотренные этапы отражают лишь главные формы обменных процессов. Как на втором, так и на третьем этапах выделяющаяся энергия накапливается в виде энергии химических связей макроэргических соединений (это вещества, имеющие хотя бы одну макроэргическую связь, напр., АТФ, ЦТФ, ТТФ, ГТФ, УТФ, АДФ, ЦДФ, …, креатинфосфат, 1,3-дифосфоглицериновая кислота). Так, энергия связи последнего фосфата в молекуле АТФ составляет около 10-12 ккал/моль.
Биологическая роль обмена веществ:
1. аккумуляция энергии при распаде химических соединений;
2. использование энергии для синтеза собственных веществ организма;
3. распад обновляемых структурных компонентов клетки;
Обмен белков
Для характеристики белкового (азотного) обмена веществ используется понятие «азотистый баланс» – это разность между азотом пищи и азотом мочи. В норме азотистый баланс равен нулю; положителен в растущем организме, во время активной регенерации, беременности; отрицателен – в старческом возрасте, при патологии, голодании.
А также имеется гамма-глутаминовый цикл (шунт), содержащий гамма-глутамилтранспептидазу (ГТТП, иначе гамма-глутамилтрансфераза – ГТТФ). Он имеет большое значение в определении функции печени, характеризуя степень усвоения АК гепатоцитами.
Определение индола и индикана в моче имеет диагностическое значение. Так, если отсутствует индол, то обезвреживающая функция печени в норме, а если при этом обнаруживается индикан, то в кишечнике активное гниение. Если же есть индол в моче, то имеется нарушение обезвреживающей функции печени.
Метаболизм аминокислот
Гидролиза белков пищи,
Общие пути обмена веществ
1. Переаминирование (открыто в 1937 г. Браунштейном и Крицмом).






