

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Топ:
Выпускная квалификационная работа: Основная часть ВКР, как правило, состоит из двух-трех глав, каждая из которых, в свою очередь...
Оснащения врачебно-сестринской бригады.
Методика измерений сопротивления растеканию тока анодного заземления: Анодный заземлитель (анод) – проводник, погруженный в электролитическую среду (грунт, раствор электролита) и подключенный к положительному...
Интересное:
Искусственное повышение поверхности территории: Варианты искусственного повышения поверхности территории необходимо выбирать на основе анализа следующих характеристик защищаемой территории...
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Подходы к решению темы фильма: Существует три основных типа исторического фильма, имеющих между собой много общего...
Дисциплины:
|
из
5.00
|
Заказать работу |
Есть разные типы жировой ткани
Жировая ткань представляет собой совокупность клеток адипоцитов (лат. adeps - жир, и kytos - полый пузырек), главной функцией которых является запасание энергии в виде жира. В состав структуры жировой ткани, наряду с адипоцитами и преадипоцитами, входят фибробласты, тучные клетки, клетки нервной ткани и эндотелия, макрофаги.
Выделяют два основных типа жировой ткани - белая и бурая, которые отличаются друг от друга по выполняемым функциям, роли в организме и предназначению. Белая жировая ткань имеет белый или желтоватый цвет, в то время как бурая жировая ткань имеет действительно бурый, коричневатый цвет, который обусловлен высокой васкуляризацией и большим количеством митохондрий с находящимися в них цитохромами. У человека белой жировой ткани гораздо больше, чем бурой. В организме здорового взрослого на нее приходится около 15% общей массы тела. В настоящее время постулировано также существование бежевой жировой ткани.
Жировая ткань распределяется в организме неравномерно. Выделяют два типа расположения: центральный (абдоминальный, андроидный, тип яблока) и периферический (гиноидный, тип груши). При центральном типе ожирения жир откладывается главным образом в брюшной полости, при периферическом - больше под кожей. При центральном типе ожирения вокруг внутренних органов имеется больше бурого жира, при периферическом ожирении под кожей откладывается метаболически неактивный белый жир. Последствия центрального и периферического ожирения также различны. Диабет, гипертония, атеросклероз - это осложнения андроидного ожирения.
Белая жировая ткань выполняет несколько функций
Адипоциты белой жировой ткани - крупные клетки сферической формы, обычно диаметром от 30 до 70 мкм, однако эти клетки могут увеличивать свой диаметр в 20 раз, а свой объем в несколько тысяч раз.
В процессе эмбрионального развития жировая ткань развивается из мезенхимы - эмбриональной соединительной ткани. Мезенхима дает начало всем остальным клеткам соединительной ткани, в том числе и адипоцитам. Человек рождается с готовыми жировыми отложениями, которые начинают формироваться на 30 неделе внутриутробного развития.
Зрелые жировые клетки у взрослого не делятся, но клетки-предшественники адипоцитов имеются на протяжении всей жизни и в норме делятся только в двух жизненных периодах - последний триместр эмбрионального развития и пубертатный период. Однако в любом возрасте при избыточном накоплении жира в клетке (гипертрофии) и достижении критической массы преадипоциты получают сигнал и начинают делиться (гиперплазия). И если у обычного худощавого человека количество адипоцитов около 35 млрд, то при ожирении оно способно увеличиться в 4 раза. При похудении адипоциты не исчезают, а только уменьшаются в размере.
Адипоцит, как правило, почти полностью заполнен большой липидной каплей, которая составляет 65-85% массы и оттесняет митохондрии и ядро клетки на периферию, в узкое свободное от жира пространство вблизи плазматической мембраны. Липидная капля покрыта мембраной, состоящей из монослоя фосфолипидов, жирнокислотные остатки которых погружены внутрь, а также из встроенных в монослой и покрывающих его белков (перилипин и адипофилин). Основную часть липидов составляют триацилглицеролы, присутствуют также холестерол (ХС), диацилглицеролы (ДАГ) и свободные жирные кислоты (СЖК).
Функции
1. Термоизолирующая - развитая жировая ткань образует теплоизоляционный слой, защищающий организм от действия очень низких температур. Особенно хорошо это проявляется у животных Арктики и Антарктики (тюлени, моржи, пингвины).
2. Механическая защита органов и создание "ложи", например, "жировая подушка" почки удерживает ее на месте.
3. Пластическая функция заключается в том, что белая жировая ткань замещает ткань некоторых органов при их инволюции. Примерами являются тимус, молочные железы, костный мозг.
4. Регуляторная функция. Являясь одним из компонентов стромы костного мозга, белый жир формирует микроокружение для развивающихся форменных элементов крови и обеспечивает их питательными веществами и ростовыми факторами.
5. Резервно-энергетическая - большую часть массы адипоцитов составляют триацилглицеролы (ТАГ), являющиеся энергетическим субстратом.
6. Эндокринная функция. Жировая ткань является активным эндокринным органом, выделяющим разнообразные биологически активные молекулы.
7. Также функцией жировой ткани является депонирование жирорастворимых витаминов A, D, Е. С некоторым усилием к функции адипоцитов можно отнести связывание и накопление вредных гидрофобных соединений, и препятствование их переносу в другие ткани, т.е. пассивное обезвреживание, например, как в случае с избытком билирубина при гемолизе.
Запасание жира - липогенез
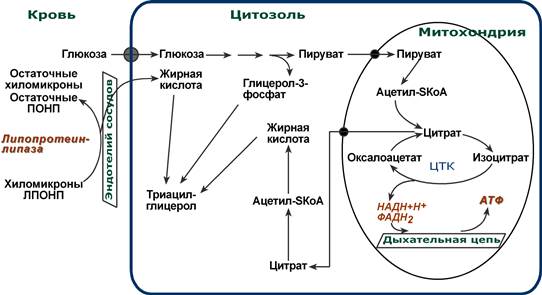
Запасы триацилглицеринов в адипоцитах пополняются за счет использования жирных кислот, поступающих из кишечного тракта после приема жирной пищи (переносятся в составе ТАГ хиломикронами) или жирных кислот, синтезированных из глюкозы в печени (переносятся ЛПОНП в составе ТАГ). Отрыв жирных кислот от ТАГ в липопротеинах крови обеспечивает липопротеинлипаза, находящаяся на эндотелии кровеносных капилляров. Адипоциты включают полученные жирные кислоты в состав триацилглицеринов и хранят в виде больших жировых капель (Подробно о хиломикронах и ЛПОНП).
При высоком потреблении углеводов жировая ткань также поглощает глюкозу, превращает их (через пируват и ацетил-СоА) в насыщенные и моненасыщенные жирные кислоты, Но в целом считается, что у человека синтез жиров из глюкозы происходит большей частью в гепатоцитах, а запасы в адипоцитах пополняются за счет уже готовых жирных кислот.
Липолиз - новое прочтение
Мобилизация жира
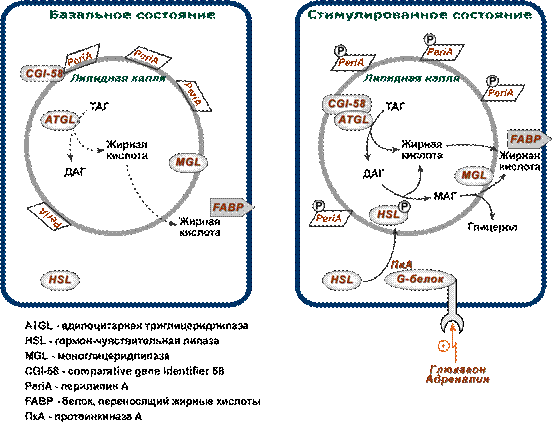
В настоящее время, после открытия в 2004 году адипоцитарной триглицеридлипазы (ATGL, adipose triglyceride lipase, деснутрин), прежняя концепция липолиза получила новое прочтение. ATGL несет ответственность за большую часть липолитической активности как в базальных, так и в стимулированных условиях.
В обычных условиях на поверхности липидной капли находятся белок перилипин, ATGL и ее коактиватор под названием CGI-58 (comparative gene identification-58, или также Abhd5, abhydrolase domain-containing 5).
Гормон-чувствительная липаза (HSL, hormone sensitive lipase) и белок, переносящий жирные кислоты (БПЖК, FABP, fatty acid binding protein) в это время преимущественно находятся в цитоплазме.
В спокойном адипоците
· ATGL отщепляет жирные кислоты (С16-С18) во 2-м положении триацилглицерола с образованием продукта 1,3-ДАГ. Происходит базальный липолиз.
В стимулированном адипоците
· перилипин, фосфорилируемый протеинкиназой А, изменяет свою конформацию и уходит от поверхности липидной капли в цитозоль.
· белок CGI-58 отделяется от перилипина и присоединяется к ATGL, формируя с ней активный комплекс и меняя ее специфичность.
· триглицеридлипаза (ATGL), после связывания с CGI-58, начинает отщеплять жирные кислоты от 1-го положения три ацилглицеролов, продуцируя 2,3-ДАГ. Именно 2,3-ДАГ является субстратом гормон-чувствительной липазы (HSL).
· гормон-чувствительная липаза (HSL) фосфорилируется протеинкиназой А и/или протеинкиназой G и в такой активной форме мигрирует к липидной капле. Отсутствие перилипина позволяет этой липазе подобраться к ядру липидной капли, где она гидролизует 2,3- ди ацилглицеролы до 2-моно ацилглицеролов и свободной жирной кислоты.
· МАГ-липаза находится как в цитозоле, так и на поверхности липидной капли и гидролизует моно ацилглицеролы.
Хотя главным субстратом HSL являются диацилглицеролы, также она может гидролизовать ТАГ и МАГ, в соотношении скоростей гидролиза 20: 2: 1 для ДАГ:ТАГ:МАГ соответственно.
Жирные кислоты покидают жировую клетку и, связываясь с сывороточным альбумином, переносятся кровотоком. Затем они освобождаются от альбумина и с помощью тканевых белков, переносящих жирные кислоты, проникают в мишеневые клетки.
Субстратный цикл
Базальный распад и синтез триацилглицеролов в жировой ткани объединены в субстратный цикл, который заключается в том, что 1,3-ДАГ, продукт малоактивной ATGL, является предпочтительным субстратом одного из ферментов синтеза триацилглицеролов – ДАГ-ацил-трансферазы. В результате ATGL и ДАГ-ацил-ТФ действуют скоординированно и поддерживают непрерывный цикл гидролиза-реэтерификации ТАГ.
В состоянии покоя около 70% жирных кислот, высвобожденных при участии малоактивной ATGL, не покидают клетку и включаются обратно в состав ТАГ. Остальные 30% жирных кислот выходят в кровь. Вышедшие в кровь жирные кислоты используются в тканях, и их часть неминуемо попадает в печень, где они реэтерифицируются в ТАГ, далее формируется их транспортная форма ЛПОНП. При метаболизме последних в плазме крови образуются остаточные ЛПОНП, некоторая часть которых превращается в синусоидах печени в атерогенные ЛПНП. Нетрудно представить, что количество образуемых ЛПОНП и ЛПНП напрямую зависит от жировой массы, что связывает ожирение и развитие атеросклероза.
Активации субстратного цикла жирных кислот также способствует часто наблюдающийся дисбаланс между потреблением насыщенных и полиненасыщенных жирных кислот, так как липидная капля в адипоците окружена монослоем фосфолипидов, которые должны содержать ПНЖК. При нарушении синтеза фосфолипидов доступ ATGL-липазы к триацилглицеролам облегчается и их гидролиз ускоряется, возрастает доля насыщенных ЖК, выходящих в кровь.
Избыток насыщенных ЖК крови легко переходит в клеточные мембраны и задерживается здесь, образуя кластеры. Это меняет свойства мембраны, нарушает ионные потоки, ухудшает взаимодействие рецепторов с их лигандами.
Лептин
Лептин - один из основных гормонов жировой ткани. Он играет ключевую роль в регуляции энергетического гомеостаза, сигнализируя головному мозгу об увеличении запаса жира в организме.
Экспрессия мРНК гена ob, кодирующего лептин, зависит от ряда факторов:
· размер адипоцито в – первостепенная составляющая в синтезе лептина, т.е. большие жировые клетки продуцируют больше лептина,
· также гормоны глюкокортикоиды, эстрогены и инсулин увеличивают синтез лептина.
Лептин взаимодействует со специфическими рецепторами в гипоталамусе (паравентрикулярными, латеральными, вентромедиальными, дорсомедиальными) и снижает. пищевое поведение через подавление экспрессии генов и биосинтеза белков, стимулирующих аппетит и пищевое поведение – нейропептида Y, AGRP-белка и меланин-концентрирующего гормона. Параллельно лептин активирует экспрессию генов α-меланоцит-стимулирующего гормона и CART (cocaine amphetamine regulated transcript) в других нейронах, отвечающих за снижение пищевого поведения.
Кроме того, лептин является гипоталамическим стимулятором симпатической нервной системы, инициатором несократительного термогенеза благодаря экспрессии UCP-1 в адипоцитах бурой жировой ткани, что увеличивает основной обмен и тепловое рассеяние.
Кроме прямых липостатических функций, лептин обладает другими эндокринными и нейроэндокринными функциями, модулируя активность тиреотропной, соматотропной, кортикотропной и гонадотропной осей, изменяя чувствительность к инсулину в скелетных мышцах и печени.
Адипонектин
Адипонектин – полипептид, секреция которого стимулируется инсулином. Адипонектин регулирует энергетический гомеостаз и оказывает антивоспалительный и антиатерогенный эффекты. В отличие от других адипокинов его концентрация снижается при ожирении и при инсулинорезистентности. Вероятно, адипонектин влияет на активность TNF-α и выполняет защитную роль при гипергликемии, инсулинорезистентности и атеросклерозе.
Резистин
Резистин синтезируется преимущественно преадипоцитами и, в меньшей степени, зрелыми адипоцитами висцеральной жировой ткани.
Действие резистина вызывает контринсулярные эффекты – прекращение инсулин-стимулированного входа глюкозы в адипоцит, развитие инсулинорезистентности и нарушение толерантности к глюкозе. Резистин играет ключевую роль в появлении избыточной массы тела у больных сахарным диабетом 2 типа.
Адипсин
Адипсин животных и человека идентичен фактору D системы комплемента и в целом он способствует проявлению эффектов инсулина на жировую ткань. Экспрессия адипсина регулируется инсулином и глюкокортикоидами.
Интерлейкин-6
Интерлейкин-6 является полифункциональным цитокином, более 30% его содержания обеспечивается жировой тканью, в основном висцеральными адипоцитами, которые образуют его в 2-3 раза больше, чем подкожные. На синтез интерлейкина-6 влияет ФНОα, глюкокортикоиды и катехоламины.
Интерлейкин-6 оказывает тормозящее влияние на адипогенез и способствует снижению секреции адипонектина. В связи со спецификой расположения висцеральной жировой ткани секретируемый ею интерлейкин-6 оказывает прямое воздействие на метаболизм веществ в печени, также снижает синтез липопротеинлипазы в жировой ткани. Этот цитокин стимулирует термогенез и активность адреналовой системы, подавляет активность гормонов половой системы, секрецию тиреотропного гормона, нарушает цикличность секреции лютеинизирующего гормона.
Висфатин
Висфатин синтезируется преимущественно в висцеральной жировой ткани. Он обладает инсулин-имитирующим действием, стимулируя транспорт глюкозы в периферические ткани и тормозя продукцию глюкозы гепатоцитами. Подобно инсулину, висфатин связывает инсулиновый рецептор и имитирует эффекты инсулина.
Отмечено повышение концентраций циркулирующего висфатина у здоровых добровольцев при гипергликемии.
Апелин
Апелин (APLN) секретируется, кроме жировой, и в других тканях – почках, мозге, сердце. Его производство адипоцитами увеличивается во время дифференцировки и стимулируется инсулином. Действие апелина, в свою очередь, угнетает секрецию инсулина.
Влияние апелина на сосудистую систему способствует образованию новых кровеносных сосудов, высвобождает NO, расслабляя стенку артерий, стимулирует сократительную способность миокарда. В мозгу апелин участвует в регуляции потребления воды, снижает секрецию вазопрессина и задействован в регуляции аппетита.
У больных ожирением с гиперинсулинизмом плазменные значения этого адипокина существенно повышены..
Эстрогены
В жировой ткани содержится фермент ароматаза (изоформа цитохрома Р450), которая преобразует тестостерон в эстрадиол. Также ароматаза представлена в других тканях, включая гонады, мозг, кровеносные сосуды, кожу, волосяные фолликулы, кости, эндометрий. Наиболее высокой ароматазной активностью отличается жировая ткань брюшной стенки.
Физиологически роль ароматазы важна в жировой ткани у женщин во время менопаузы, когда продукция эстрадиола в яичниках существенно ослабевает. У мужчин активность фермента проявляется при андрогенном типе ожирения, которое связано с накоплением жира на животе и висцерального жира.
Заболевания
Заболевания органов и систем отражаются на состоянии жировой ткани и, соответственно, на массе тела в результате изменения рациона и частоты питания, снижения или усиления аппетита. Особенно ярко проявляется проблема при онкологических заболеваниях, нарушении желудочно-кишечного тракта и гормональных проблемах. Изменение массы тела в этом случае являются вторичными - вторичное ожирение, вторичное истощение.
Размеры адипоцита зависят от гормонального фона
1. Инсулин активирует ферментные системы, ответственные за липогенез и тормозит ферментные системы ответственные за липолиз. Чем больше количество глюкозы в крови, тем больше в крови инсулина, тем больше поступает в адипоцит из крови глюкозы, глицерина и жирных кислот, тем больше образуется внутриклеточного жира. Поэтому избыточное употребление пищевых углеводов влечёт за собой увеличение количества жира в адипоцитах.
2. Еще один гормон, обеспечивающий запасание жира – пролактин (лактотропный гормон). Метаболические эффекты пролактина заключаются в повышении липолитической активности плазмы крови (активация липопротеинлипазы) и перехода жирных кислот в ткани, в усилении реакции инсулина на введение глюкозы. Особенно ярко проявляются эффекты пролактина во время беременности и после родов, когда количество пролактин-синтезирующих клеток в гипоталамусе увеличивается и зачастую это увеличение сохраняется и после родов, что часто вызывает ожирение.
Секреция пролактина подвержена многим регуляторным влияниям. Например, при обезвоживании или гипонатриемии происходит активация системы РААС и образуемый ангиотензин II стимулирует секрецию пролактина. Стимулятором секреции пролактина также является тиреотропин-рилизинг-фактор (тиреолиберин) гипоталамуса. При первичной недостаточности тиреоидных гормонов по механизму обратной отрицательной связи повышается синтез этого фактора в гипоталамусе и происходит (даже вне беременности) повышенная секреция пролактина в кровь. Кроме ожирения, следствием такого явления могут быть галакторея, отсутствие овуляции, аменорея и бесплодие.
3. Гормоны катехоламины: адреналин и норадреналин активируют как липогенез, так и липолиз в адипоцитах. Свое влияние на эти два противоположных процесса катехоламины реализуют через разные адренорецепторы адипоцитов – α2-адренорецепторы и β-адренорецепторы. Действие через α2-адренорецепторы активирует в адипоците липогенез, влияние через β-адренорецепторы – липолиз. α2-Адренорецепторы имеют б о льш е е сродство к катехоламинам, поэтому соединение катехоламинов с β-адренорецепторами происходит только тогда, когда свободных α2-адренорецепторов уже нет. Иными словами, когда концентрация катехоламинов низка, они соединяются с α2-адренорецепторами, поддерживая в адипоците липогенез. При увеличении концентрации (острый стресс) они связываются с β-адренорецепторами, активизируя в адипоците липолиз.
4. Глюкокортикоиды изменяют соотношение α2- и β-адренорецепторов на поверхности адипоцитов.
Количество α2- и β-адренорецепторов на адипоцитах разных частей тела неодинаково, поэтому жир откладывается по телу неравномерно. Это определено индивидуальной (генетической, наследственной) особенностью и половой принадлежностью. Если количество α2-адренорецепторов преобладает над количеством β-адренорецепторов, адипоцит больше работает на запасание ТАГ. У женщин α2-адренорецепторов больше на жировых клетках в области ягодиц, бедер, коленей, у мужчин - в области подкожной жировой клетчатки живота и висцерального жира. Адипоциты в этих местах гораздо активнее накапливают жир и существенно медленнее отдают жир при похудании. Т.е. при ожирении триацилглицеролы накапливаются именно в указанных зонах, а при снижении веса тела, например, при диетах или голодании, исчезновение жира происходит в других участках тела. Каждое последующее увеличение/уменьшение массы жировой ткани будет увеличивать жировые отложения именно в этих вышеуказанных местах.
Под влиянием избыточного количества глюкокортикоидов в подкожных адипоцитах лица и верхней части туловища соотношение адренорецепторов смещается в сторону α2-адренорецепторов. Как результат, при обычных концентрациях катехоламинов ТАГ начинают наиболее интенсивно откладываться в области лица и верхней части туловища (например, при болезни Кушинга). Такое ожирение усиливается за счет сопутствующей гипергликемии и дополнительной секреции инсулина, стимулирующего липогенез.
Еще один эффект глюкокортикоидов заключается в индукции синтеза гормон-чувствительной липазы, ключевого фермента липолиза. Однако необходимо помнить, что кортизол сам по себе не стимулирует липолиз, без участия других липолитических гормонов данный эффект не проявляется.
5. Глюкагон активирует в адипоцитах ферментные системы, ответственные за липолиз, и тормозит липогенез. Количество глюкагона в крови увеличивается при голодании, а также при повышенном расходовании глюкозы с появлением гипогликемии, например, при физической нагрузке. Поэтому количество жира в жировой ткани можно уменьшить в результате голодания и физической нагрузки.
6. Гормоны щитовидной железы повышают активность Na + /К + -АТФазы и повышают потребность клетки в энергии, что решается ускорением катаболизма. Поскольку на обеспечение работы этого фермента (поддержание мембранного потенциала) затрачивается от 40 до 60% всей клеточной АТФ, малейшая его активация резко повышает катаболизм гликогена и триацилглицеролов для восполнения затрат АТФ. Параллельно тиреоидные гормоны снижают активность фосфодиэстеразы, гидролизующей цАМФ и уменьшающей катаболизм. Таким образом, избыточное действие тиреоидных гормонов приводит к снижению массы тела. Общий катаболический эффект тироксина и трийодтиронина присутствует во всех тканях, кроме печени.
Способностью активировать в адипоцитах процесс липолиза обладают также гормоны СТГ, АКТГ, тестостерон.
7. Грелин - пептид, вырабатываемый в пустом желудке. Оказывая влияние на гипоталамус и гипофиз, грелин вызывает чувство голода. Он участвует в адаптивном ответе на снижение веса, т.е. в ответ на голодание и потерю веса происходит увеличение его концентрации в крови и повышенное потребление пищи. По всем эффектам грелин и лептин оказывают реципрокное действие, реализуемое на периферии и на уровне центральных гипоталамических механизмов. При этом такие взаимоотношения присутствуют как на этапе кратковременной регуляции потребления пищи, так и долговременной регуляции энергетического гомеостаза.
Одновременно грелин стимулирует такие разные процессы как секреция гормона роста и формирование новых синапсов в головном мозге, т.е. усиливает процессы запоминания. Как тут не вспомнить поговорку: "Сытое брюхо к учению глухо".
8. Гормон адипоцитов лептин, попадая с током крови в гипоталамус, уменьшает образование в гипоталамусе нейропептида Y, который стимулирует пищевое поведение. Адипоцит начинает синтезировать и секретировать в кровь лептин при увеличении количества внутриклеточных ТАГ. Иными словами при увеличении размеров адипоцита количество лептина возрастает и, через нейропептид Y, снижается потребление пищи. Однако существует феномен снижения чувствительности рецепторов гипоталамуса к лептину, также встречается генетическое изменение структуры лептина. Из-за этого стимуляция лептином гипоталамуса не достаточна и синтез нейропептида Y не прекращается при увеличении размера адипоцита, поиск и потребление пищи продолжаются.
Мы есть то, что мы едим
Накопление жировой массы ускоряется:
· при избытке субстрата для синтетических реакций, а именно жира и углеводов,
· при низкой физической активности.
Абсолютно необходимым условием увеличения массы тела являются повышение синтеза подкожного жира или снижение его распада. Вопрос баланса энергии - ключевой при рассмотрении проблемы ожирения. По одному шутливому выражению, ожирение - это синдром двойного невставания, т.е. отказ больного вовремя встать из-за стола и нежелание встать с дивана ради физической нагрузки.
Наличие в продуктах избыточного количества жира обеспечивает быстрый прирост объема адипоцитов и жировой массы.
При этом нежелательной особенностью является более приятный вкус у жирных продуктов. Подобные вкусовые нюансы закреплены эволюционно для повышения шансов на выживание при голодании. Ведь если индивид отдавал предпочтение жирной пище, то и запасов энергии у него было больше. К сожалению, пищевая промышленность знает об этом и активно использует вкусовые предпочтения для продвижения своей продукции, добавляя "скрытый" жир в кондитерские, молочные и другие изделия.
Избыток углеводов в продуктах тоже неблагоприятно сказывается на состоянии жировой массы, хотя и не так быстро и явно, как чистый жир. Жировая ткань превращает в ТАГ лишь 20% избыточной глюкозы. И считается, что остальные 80% лишнего сахара сгорают в печени или мышцах, повышая величину основного обмена. Только единоразовый прием около 100 г углеводов (особенно при потреблении моно- и дисахаридов) приводит в печени к превращению глюкозы в нейтральный жир с последующим его переносом в адипоциты.
При психическом нарушении как нейрогенная булимия больной поглощает огромное количество продуктов, даже не относящихся к вкусной деликатесной пище. Обычно такое состояние наступает после длительной диеты с целью похудеть. Рекордное потребление энергии при таком расстройстве составило 50000 ккал/сутки.
Необходимо заметить, что избыток субстрата, особенно у девочек и женщин, в определенные возрастные периоды вызывает стойкое ожирение. Объясняется это способностью жировых клеток к делению, причем деление происходит в зависимости не столько от генетических программ, сколько от количества жира в клетке. Переполнение жировых клеток у плода в последние 3 месяца беременности, в раннем младенчестве, пубертатном и преклимактерическом возрасте стимулирует эти клетки к делению. Жировая ткань разрастается, повышается ее способность к запасанию и синтезу жира.
Пониженная физическая активность в развитых странах является одной из существенных причин проблемы ожирения. Дело в том, что мышцы, как при работе, так и в спокойном состоянии сжигают огромное количество глюкозы и жирных кислот. Уменьшение доли используемых мышцей субстратов приводит к перенаправлению их избытка в жировую ткань.
Холостые циклы
Бесполезные или " холостые " циклы – это активное протекание противоположных реакций, требующих затрат энергии, например, реакции синтеза и распада глюкозо-6-фосфата, фруктозо-1,6-дифосфата, цикл липолиз-липогенез.
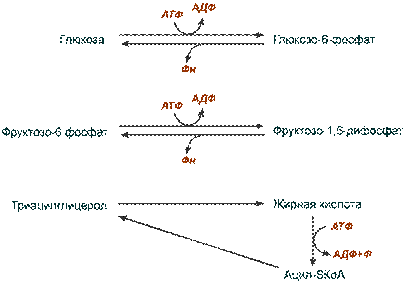
Примеры холостых циклов
В результате таких циклов возрастает потребность клетки в энергии АТФ, которая черпается из окисления глюкозы или жирных кислот. Таким образом, расход энергетических субстратов возрастает, скорость катаболизма увеличивается.
Повышение активности Na + /К + -АТФазы под действием тиреоидных гормонов также можно отнести к "холостым" циклам, так как величина мембранного потенциала существенно не изменяется из-за обратной диффузии электролитов по градиенту концентрации, а АТФ расходуется гораздо больше.
Бурая жировая ткань
Строение
Клетки белой и бурой жировой ткани значительно отличаются друг от друга. Клетка бурой жировой ткани имеет по всей цитоплазме много мелких жировых капель, окружающих большое количество митохондрий, обильно снабжена окончаниями симпатических нервов и кровеносными сосудами. В эмбриогенезе она развивается из миобластов миотома сомитов, т.е. из предшественников скелетной мускулатуры, а не из предшественников соединительной ткани, подобно белым адипоцитам.
Бурая жировая ткань у младенцев составляет около 5% от массы их тела. Она расположена подкожно, между лопатками, в виде небольших отложений вокруг мышц и кровеносных сосудов шеи, в щеках (комочки Биша), а также в большом количестве – в подмышечных впадинах. Более глубокие отложения находятся в переднем средостении, вдоль позвоночника, вокруг почек, надпочечников, аорты, между лопатками, около почек и щитовидной железы. Между лопатками, на грудной клетке и на плечах у младенцев бурые адипоциты граничат с белыми, но нет четкой грани между клетками, среди бурых адипоцитов на "территории" бурой жировой ткани располагаются в некотором количестве белые адипоциты.
По мере взросления количество клеток бурой жировой ткани в указанных зонах снижается, но совсем не исчезает. По частоте встречаемости в организме одна клетка бурого жира, в среднем, приходится на 100-200 белых адипоцитов.
Функции
Функцией бурой жировой ткани является оперативное обеспечение организма тепловой энергией (адаптивный термогенез). Основной механизм термогенеза определяется тем, что в митохондриях клеток бурых адипоцитов процессы окисления и фосфорилирования не сопряжены (разобщены) и при окислении поступающих сюда жирных кислот происходит, в основном, выделение тепла, а не синтез АТФ.
Разобщение окисления и фосфорилирования обеспечивают специальные белки – термогенины (разобщающие белки, uncoupling proteins, UCP). Существуют несколько изоформ разобщающих белков: UCP-1 находится преимущественно в бурой жировой ткани, UCP-2 – в бурой и белой, UCP-3 – в cкелетных мышцах, UCP-4 и UCP-5 – преимущественно в мозге.
Адаптивный термогенез – это повышение теплопродукции в ответ:
· на изменение окружающей среды (снижение температуры) и на переохлаждение организма. При этом активируется симпатическая нервная система и стимулирует липолиз в бурых адипоцитах,
· на стимуляцию катехоламинами через β3-адренорецепторы – повышение транскрипции гена UCP-1, что приводит к увеличению энергозатрат,
· на избыток потребления пищи или изменение ее состава (поступление жиров),
· на секрецию лептина (усиление транскрипции гена UCP-1),
· на какие-то патологические воздействия.
У человека адаптивный термогенез ярко выражен у новорожденных, но при взрослении становится менее востребованным, заменяясь на иные способы сохранения тепла.
У взрослых отсутствие (резкое снижение) бурой жировой ткани встречается в 10% всех случаев ожирения.
Бурой жировой ткани много у животных, которые впадают в зимний сон и зимнюю спячку, температура тела у них поддерживается за счет адаптивного термогенеза.
Небольшие количества бурой жировой ткани у здоровых взрослых людей осуществляют постпрандиальный (возникающий после еды) термогенез, что в определенной степени препятствует отложению жира на фоне избытка пищи. При ожирении клетки бурой жировой ткани практически отсутствуют и, соответственно, процесс постпрандиального термогенеза не происходит.
Бежевая жировая ткань
Установлено, что помимо белой и бурой жировых тканей имеется промежуточная разновидность – бежевая жировая ткань. Эта ткань является промежуточной между белой и бурой жировыми тканями. Бежевые адипоциты первоначально имеют крайне низкое количество термогенина, но, как и классические бурые адипоциты, реагируют на определенную стимуляцию (понижение температуры, мышечная работа) началом высокой экспрессии термогенина и превращением в активные теплообразующие адипоциты.
Некоторыми авторами возможность перехода белой жировой ткани в бурую жировую ткань через бежевую называется доказанным фактом.
По другим авторам, бежевая жировая ткань является самостоятельной тканью, возникшей из белой и приблизившейся к бурой по морфологическим и биохимическим показателям. Но, тем не менее, она выделяется как отдельная ткань.
Независимо от точек зрения, развитие и наличие бежевой жировой ткани служит важным механизмом поддержания энергетического и температурного гомеостаза.
При работе скелетных мышц в них повышается содержание особого белка - фактора транскрипции PGC-1a (peroxisome proliferator-activated receptor Gamma Coactivator 1-alpha), этот белок также экспрессируется в бурой жировой ткани, сердце и почках. Он стимулирует увеличение количества другого белка FNDC5 (fibronectin type III domain-containing protein 5).
У белка FNDC5 имеется две функции:
1. Даже в очень низких концентрациях он увеличивает синтез мРНК белка термогенина в 70-100 раз.
2. При его расщеплении образуется секретируемый мышцами полипептид, который был назван ирисином. Этот гормон является посредником между физическими нагрузками и различными тканями организма, в том числе белой и бурой жировыми тканями.
Вырабатываясь в работающих мышцах, ирисин транспортируется по крови в белые адипоциты, меняет акценты транскрипции и преобразует белую жировую ткань в бежевую.
Ирисин вырабатывается мышечной тканью не только при физической работе, но и при действии на организм холода. Так, при дрожании от холода в течение 10-15 минут вырабатывается такое же количество ирисина, как и при часовой езде на велосипеде со средней интенсивностью.
Название ирисин дано по имени древнегреческой богини Ириды, которая являлась богиней радуги и вестницей богов Зевса и Геры.
Есть разные типы жировой ткани
Жировая ткань представляет собой совокупность клеток адипоцитов (лат. adeps - жир, и kytos - полый пузырек), главной функцией которых является запасание энергии в виде жира. В состав структуры жировой ткани, наряду с адипоцитами и преадипоцитами, входят фибробласты, тучные клетки, клетки нервной ткани и эндотелия, макрофаги.
Выделяют два основных типа жировой ткани - белая и бурая, которые отличаются друг от друга по выполняемым функциям, роли в организме и предназначению. Белая жировая ткань имеет белый или желтоватый цвет, в то время как бурая жировая ткань имеет действительно бурый, коричневатый цвет, который обусловлен высокой васкуляризацией и большим количеством митохондрий с находящимися в них цитохромами. У человека белой жировой ткани гораздо больше, чем бурой. В организме здорового взрослого на нее приходится около 15% общей массы тела. В настоящее время постулировано также существование бежевой жировой ткани.
Жировая ткань распределяется в организме неравномерно. Выделяют два типа расположения: центральный (абдоминальный, андроидный, тип яблока) и периферический (гиноидный, тип груши). При центральном типе ожирения жир откладывается главным образом в брюшной полости, при периферическом - больше под кожей. При центральном типе ожирения вокруг внутренних органов имеется больше бурого жира, при периферическом ожирении под кожей откладывается метаболически неактивный белый жир. Последствия центрального и периферического ожирения также различны. Диабет, гипертония, атеросклероз - это осложнения андроидного ожирения.

Индивидуальные очистные сооружения: К классу индивидуальных очистных сооружений относят сооружения, пропускная способность которых...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Археология об основании Рима: Новые раскопки проясняют и такой острый дискуссионный вопрос, как дата самого возникновения Рима...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!