ugra (грозный), babhru (красновато-коричневый; см. русск. бобр) - постоянный эпитет Рудры. Также babhru (коричневый) - конская масть, цвет сомы, Агни и игральных костей, сделанных из лесных орехов
1 Это поклонение (тому) грозному, коричневому (ugraya babhrave),
Кто среди игральных костей властвует над телом.
Жиром я хочу одарить Кали.
Да будет он милостив к нам в таком (положении)!
2 О Агни, отвези ты жир для (нимф-русалок) апсарас.
Пыль для игральных костей, песок и воду!
Наслаждаясь жертвенным даром в соответствии со своей долей,
Боги радуются жертвам обоего рода.
3 Апсарас пируют на общем пиру
Между жертвенной повозкой и солнцем.
Пусть они смешают с жиром мои руки!
Пусть отдадут мне во власть соперника-игрока!
4 Неудачу — противнику в игре!
Нас полей жиром!
Как дерево — молнией срази (того),
Кто играет против нас!
5 Кто создал это богатство для нашей игры.
Кто (создал) взятку и остаток игральных костей,
Тот бог, наслаждаясь этим нашим возлиянием, —
Пусть пируем мы на общем пиру вместе с гандхарвами!
6 «Живущие вместе» — вот ваше название.
Ведь игральные кости — (те), кто видит грозного:
они несут власть.
Вас таких, о капли, мы хотим почтить возлиянием!
Пусть будем мы господами богатств!
7 Если я, умоляя, призываю богов.
Если жили мы как ведийские ученики,
Если я выхватываю коричневые игральные кости,
Да будут они милостивы к нам в таком (положении)!
...счастливые взятки при выхватывании костей (glahe krtani krnvanаm)... — Требовалось, чтобы из кучи игральных костей игрок выхватил какое-то их количество. Если их число делилось на 4 без остатка, то это была «счастливая взятка» (krta-)
...кроме счастливой взятки, делящейся на четыре без остатка, есть tretа-, когда остается три кости; dvapara-, когда остается две кости, и kali-, когда остается одна кость, несчастливая взятка.
- АВ. IV, 38. На удачу при игре в кости - с помощью апсарас (русалок)
Ригведа. Мандалы I-IV (перевод Т.Я. Елизаренковой) 19.6Мб
Ригведа. Мандалы V-VIII (перевод Т.Я. Елизаренковой) 8.7Мб
Ригведа. Мандалы IX-X (перевод Т.Я. Елизаренковой) 13.4Мб
Атхарваведа. Избранное (Перевод, комментарии и вступ. статья Т.Я. Елизаренковой) 5.7Мб
http://www.bolesmir.ru/index.php?content=text&name=o285

Общий план строения иммуноглобулинов: 1) Fab; 2) Fc; 3) тяжёлая цепь; 4) лёгкая цепь; 5) антиген-связывающийся участок; 6) шарнирный участок - см. Антитела
Как наша иммунная система понимает устройство антигена и подбирает подходящее для него антитело?
Рассмотрим этот процесс на примере развития бактериальной инфекции. Например, вы поцарапали палец. При повреждении кожи в рану чаще всего попадают бактерии. При повреждении любой ткани организма сразу же запускается воспалительная реакция. Поврежденные клетки выделяют большое количество разных веществ — цитокинов, к которым очень чувствительны нейтрофилы и макрофаги. Реагируя на цитокины, они проникают через стенки капилляров, «приплывают» к месту повреждения и начинают поглощать и переваривать попавших в рану бактерий — так запускается неспецифический иммунитет, но до синтеза антител дело пока еще не дошло.
Расправляясь с бактериями, макрофаги выводят на свою поверхность разные их кусочки, чтобы познакомить Т-хелперов и B-лимфоцитов со строением этих бактерий. Этот процесс называется презентацией антигена. Т-хелпер и B-лимфоцит изучают кусочки переваренной бактерии и подбирают соответствующую структуру антитела так, чтобы потом оно хорошо «прилипало» к таким же бактериям. Так запускается специфический гуморальный иммунитет. Это довольно длительный процесс, поэтому при первом контакте с инфекцией организму может понадобиться до двух недель, чтобы подобрать структуру и начать синтезировать нужные антитела.
После этого успешно справившийся с задачей B-лимфоцит превращается в плазматическую клетку и начинает в большом количестве синтезировать антитела. Они поступают в кровь, разносятся по всему организму и связываются со всеми проникшими бактериями, вызывая их гибель. Кроме того, бактерии с прилипшими антителами гораздо быстрее поглощаются макрофагами, что также способствует уничтожению инфекции...
Как устроен иммунитет: Объясняем по пунктам. 11 октября 2018
https://yandex.ru/health/turbo/articles?id=3387
Анализ индивидуальных репертуаров Т-клеточных рецепторов
В организме взрослого человека насчитывается порядка триллиона Т-лимфоцитов, задача которых — защитить нас от инфекций и онкологических заболеваний. Задача эта невероятно сложная, так как различных вирусов, бактерий и других паразитов очень много, равно как и мутаций, которые могут привести к развитию онкологических заболеваний. Кроме того, паразиты умело эволюционируют, меняют хозяев — постоянно появляются новые угрозы. Таким образом, заложить в наш организм защитную реакцию к каждому отдельному патогену оказывается невозможно, да и не хватило бы для этого емкости генома. Здесь мы расскажем о том, как адаптивный иммунитет решает эту непростую задачу, а также о том, как мы можем за этим наблюдать.
Адаптивный иммунитет решает свою задачу — противостояние широкому и изменчивому спектру патогенов вокруг и внутри нас — пожалуй, наиболее изящным способом. Он генерирует огромное разнообразие случайных последовательностей, кодирующих антитела и Т-клеточные рецепторы (T cell receptors, TCR), уникальные (точнее, почти уникальные) для каждого клона B- и Т-клеток соответственно.

Рисунок 1. В каждом из нас — триллион Т-лимфоцитов. У них разные функции, но каждый их них несет на поверхности молекулы своего Т-клеточного рецептора, способного распознать «свою» молекулу антигена в составе комплекса MHC и защитить нас от инфекции или онкологии.
...До того как Т-лимфоцит узнает молекулу патогена, он называется наивным Т-лимфоцитом. Такие наивные Т-лимфоциты чрезвычайно разнообразны: по современным оценкам, у каждого человека можно насчитать более 100 миллионов уникальных вариантов TCR.
Может показаться парадоксальным, но большая часть этого колоссального разнообразия TCR никогда в жизни нам не понадобится. Однако именно это разнообразие является залогом того, что для новой инфекции или опухоли найдутся специфичные Т-лимфоциты, распознающие антигены, характерные для данного патогена или патологических клеток. Здесь можно провести аналогию с огромной коллекцией ключей, из которых для (почти) любой новой двери найдется подходящий вариант.
Таким образом, разнообразие TCR для каждого организма в значительной степени определяет, сколько разных антигенов он способен распознать. Чем оно выше, тем выше шансы выработать эффективный иммунный ответ при встрече с новым патогеном.
Наивные Т-лимфоциты, распознавшие свой «замок» — антиген, — становятся эффекторными. Они активно размножаются и атакуют зараженные или злокачественные клетки, а также инструктируют другие клетки иммунной системы.
В то же время Т-клетки памяти — клональные копии эффекторных Т-лимфоцитов, участвовавших в иммунном ответе — сохраняют высокую численность на многие годы, а то и на десятки лет. Они защищают нас от повторного заболевания, наряду и во взаимодействии с B-лимфоцитами, производящими антиген-специфичные антитела и также образующими клональные популяции клеток памяти. Именно на феномене памяти адаптивного иммунитета основана практика вакцинации.
1. Формирование исходного репертуара TCR на этапе рекомбинации — из какого сора...
Огромное разнообразие вариантов Т-клеточных рецепторов, как и разнообразие антител, формируется в результате относительно случайной «сборки» — событий рекомбинации, в ходе которых из имеющегося в геноме набора так называемых V-, D-, и J-сегментов выбирается по одному варианту.
Произвольная комбинация этих сегментов собирается в новый ген, а на стыке сегментов — в местах «сварки» — происходит дополнительное случайное удаление и добавление нуклеотидов. Таким образом, место стыка сегментов V и J (D-сегмент совсем короткий, и его фактически «заваривает» в середину) характеризуется наибольшей гетерогенностью. И, как правило, именно этот гипервариабельный участок в дальнейшем играет ключевую роль в распознавании антигена — является основной «бородкой» «ключа».
Однако после первичной сборки TCR большая часть (95–98%) Т-лимфоцитов погибает в тимусе в результате позитивной и негативной селекции, направленной на то, чтобы отобрать из множества случайно генерируемых последовательностей TCR работоспособные и безопасные для организма варианты. Эту жесткую селекцию пройдут только те TCR, которые, с одной стороны, принципиально способны распознавать антигены в контексте MHC, а с другой — не проявляют сильного взаимодействия с MHC, несущими собственные пептиды организма. Такой отбор необходим для того, чтобы избежать аутоиммунных реакций — атаки Т-клеток на собственные клетки хозяина.
Интересно, что существенная часть зрелых Т-лимфоцитов помимо генов функциональных TCR несет на второй (гомологичной) хромосоме также и последовательности нефункциональных цепей TCR со сбитой рамкой считывания либо стоп-кодоном, нарушающими синтез белка. Такие TCR не экспрессируются на поверхности клеток, а значит и не проходят позитивную и негативную селекцию в тимусе. Таким образом, эти бесполезные на первый взгляд последовательности могут нам многое рассказать о процессе рекомбинационных событий, в ходе которых рождается пре-селекционный репертуар TCR [13–17]. Сюда можно отнести и частоты используемых генных сегментов, из которых происходит не вполне случайная сборка TCR, и число случайно добавленных и «отгрызенных» нуклеотидов в местах соединения сегментов, и некоторые другие характеристики, определяющие исходную структуру репертуара TCR для каждого человека и в популяции.
На основе данных по репертуарам нефункциональных вариантов TCR была построена вероятностная модель сборки [16] и установлено нуклеотидное разнообразие уже для рабочих, функциональных вариантов пре-селекционного репертуара β-цепей TCR, составившее у людей (как популяции) порядка 10^13, что существенно ниже, чем предсказания более простых моделей, считающих процессы выбора сегментов и каждый случайно вставленный нуклеотид независимыми событиями [13] (рис. 3). Создание такой модели позволяет исследователям генерировать искусственные репертуары TCR на компьютере, практически точно копируя поведение реальной рекомбинационной машинерии человека.
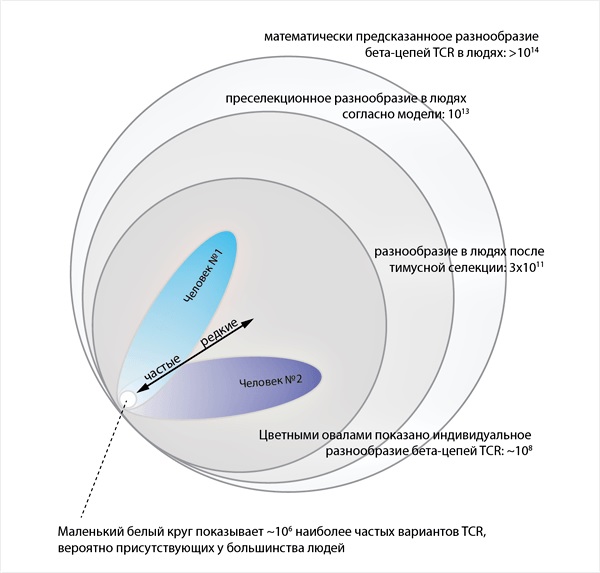
Рисунок 3. Популяционное и индивидуальное разнообразие β-цепей TCR человека
Интересно, что пре-селекционный репертуар TCR однояйцевых (генетически идентичных) близнецов характеризуется практически идентичными частотами выбора генных сегментов [17], что указывает на решающее влияние индивидуальных генетических особенностей на статистику событий рекомбинации. В то же время, например, в парах мама/свой ребенок основные характеристики пре-селекционного репертуара TCR не ближе, чем таковые для пар мама/чужой ребенок [15]...
Дмитрий Чудаков. Анализ индивидуальных репертуаров Т-клеточных рецепторов. 11 дек. 2015
https://biomolecula.ru/articles/analiz-individualnykh-repertuarov-t-kletochnykh-retseptorov
Иммунная система, генетика и происхождение видов
«Существуют два типа различий между родственными друг другу формами. Одни появляются при развитии сравнительно поздно и обусловливаются отдельными элементами генома; другие возникают при развитии с самого начала и, являясь характерными для различных родов, сводимы, вероятно, к общей структуре белков протоплазмы и ядра половых клеток, взятых в целом, или к плазмону, неразложимому, в отличие от генома, на отдельные элементы» (Филипченко, 1930)

Атхарваведа VII, 109. О корове Атхарвана
1 Кто, наслаждаясь дружбой с Брихаспати,
Создаст по своему желанию (ее) форму —
Пёструю дойную корову, данную Варуной
Атхарвану, легкодоящуюся, всегда с теленком?
V. 11. Разговор Варуны и Атхарвана
(Варуна?):
1 Как ты говорил здесь великому Асуре?
Как золотистому отцу - (ты) со сверкающим мужеством?
Дав пеструю (корову), о Варуна, как плату за жертву,
Ты, о возвращающий дар, мыслью крепко задумался. -
(Варуна):
2 Не ради удовольствия я становлюсь возвращающим дар:
Чтобы рассмотреть эту пеструю (корову), я пригоняю ее.
Благодаря какой поэзии, о Атхарван, ты сейчас (поэт)?
Благодаря какому рожденному ты знаток (всех) рожденных?
(Варуна):
3 Поистине, я глубок благодаря поэзии,
Поистине, благодаря рожденному я знаток (всех) рожденных.
Ни даса, ни арья (своим) величием
Не изменил обета, которого я буду придерживаться. -
(Атхарван):
4 Нет другого поэта, лучшего, чем ты,
Нет более мудрого (своим) пониманием, о Варуна самовластный.
Ты знаешь все эти существа.
Теперь даже этот коварный человек боится тебя.
(Атхарван):
5 Раз уж, Варуна самовластный,
Ты знаешь все рождения, о правильно ведущий,
Если ли что-нибудь другое за пределами небосвода?
Что там за пределами следующее, о безошибочный? -
(Варуна):
6 Есть одна иная (вещь) за пределами небосвода,
А за пределами одной вещи (находится то, что) труднодостижимо у нас.
Я, Варуна, зная это, провозглашаю тебе.
Пусть будут пани потерявшими почву из-под ног (?)!
Пусть уползут дасы вниз в землю! -
(Атхарван):
7 Ведь ты, Варуна, высказываешь
Много упреков в отношении возвращающих дар.
Смотри не превзойди такого рода пани!
Пусть не назовут тебя люди корыстным! -
(Варуна):
8 Пусть не назовут меня люди корыстным!
Я отдаю тебе обратно, о певец, пеструю (корову).
Охотно приходи на всякое восхваление меня
Во всех местах, обитаемых людьми! -
(Атхарван):
9 Пусть возвышенные восхваления тебя приходят
Во все места, обитаемые людьми!
Дай мне сейчас, что ты мне не дал!
Ты связанный со мною о семи шагах!
(Атхарван):
10 Общее у нас двоих происхождение, о Варуна, общий род. -
(Варуна):
Знаю я то, что у нас двоих этот род общий.
Я даю то, что тебе не дал.
Я связанный с тобою друг о семи шагах!
(Варуна):
11 Бог, наделяющий воспевающего бога жизненной силой,
Вдохновенный (поэт) большой мудрости - вдохновенного (поэта). -
(Атхарван):
Так как ты, о Варуна самовластный,
Породил Атхарвана-отца - связь богов,
Сделай ему прекрасно прославленный дар -
Ты друг наш и высшая связь!
АтхарваВеда (atharvaveda «веда заклинаний», или «веда [жреца огня] Атхарвана»)...Первоначальное название АВ не включало в себя слова «веда» (поскольку ведой этот памятник тогда не считался). Это было Атхарва или Атхарвангираса (atharvangirasa) — название, состоящее из имен двух мифических родов: Атхарвана и Ангираса (angiras — название полубожественных существ, мудрецов-риши, посредников между богами и людьми или angirasa «связанная с Ангирасами»). По индийской традиции считается, что связь АВ с двумя родами — Атхарванами и Ангирасами — отражает двоякую природу заговоров этого собрания. Белую магию (заговоры, обращенные на достижение добра) связывают с Атхарванами, а черную магию (заговоры, обращенные на достижение зла) связывают с Ангирасами.
Атхарваведа. Избранное (Перевод, комментарии и вступ. статья Т.Я. Елизаренковой) 5.7Мб
http://www.bolesmir.ru/index.php?content=text&name=o285

Ф.Г. Добржанский (слева) и Ю.А. Филипченко (справа)
...В Положениях к диссертации 1912г. Филипченко сформулировал свои убеждения: «…Процесс эволюции организмов не объясним ни так называемыми факторами Ламарка, ни подбором, а является одной из коренных особенностей живых существ. … Допуская возможность сведения отдельных жизненных процессов к чисто механическим причинам, мы едва ли будем в состоянии объяснить последними жизнь какого-либо организма в ея целом» (Фонд 813...). То есть он сделал выбор в пользу автогенеза Бэра (но не эктогенеза Ламарка или Дарвина), в пользу системного подхода Бэра (и против механизма и витализма). А в 1924г. он издал в своих переводах с немецкого и латыни наиболее важные фрагменты и работы Карла Бэра (Бэр, 1924).
В автогенезе Филипченко основа всех его научных предпочтений. Изучая гибриды диких и домашних форм в заповеднике АсканияНова, он сосредоточился на количественных признаках с полимерным наследованием, ибо эти нормальные признаки живущих в природе видов составляют эволюционно значимую часть изменчивости. В нем же основа его нелюбви к дрозофильной генетике в духе Т. Моргана, хотя заниматься ею сотрудникам и ученикам он позволял.
Каким образом Добржанский, генетик-дрозофилист, явный морганист, смог оказаться у Филипченко, скептически относившегося к генетике дрозофилы и к «папе Моргану»?
Быть может, дело в систематических работах на божьих коровках (с 1917 г.) или в занятиях Добржанского множественным действием гена на дрозофиле (его первая работа в области генетики (Dobzhansky, 1927))? (А Филипченко был склонен видеть в плейотропии основу для отмены простой схемы «один ген – один признак» и искать в ней опоры идеи плазмона).
«Подводная часть айсберга», заявленная в Положениях к диссертации, вышла на поверхность в утверждении Филипченко о необходимости различать «микро-» и «макроэволюцию» как самостоятельные области научного исследования, в пренебрежении микро- и предпочтении макроэволюции. он утверждал, что признаки высших систематических категорий определяются не генами, а чем-то иным, недостаточно выясненным, даже смутным, что он, вслед за К. Корренсом и Ф. Ветштейном, именовал «плазмоном». Последнюю статью, напечатанную в 8-м выпуске «Известий Бюро» следом за извещением о кончине Филипченко 19 мая 1930 г., он завершил своего рода завещанием:
«Существуют два типа различий между родственными друг другу формами. Одни появляются при развитии сравнительно поздно и обусловливаются отдельными элементами генома; другие возникают при развитии с самого начала и, являясь характерными для различных родов, сводимы, вероятно, к общей структуре белков протоплазмы и ядра половых клеток, взятых в целом, или к плазмону, неразложимому, в отличие от генома, на отдельные элементы» (Филипченко, 1930).
Вот этот плазмон, реален ли он? – тема, живо обсуждавшаяся в переписке Филипченко и Добржанского, энтузиаста и скептика.
Как получилось, что сотрудник Филипченко, приверженца макроэволюции, посмел заняться теорией микроэволюции и внести в нее выдающийся вклад? Но Добржанский ценил не только Филипченко, но и Дарвина. В «Происхождении…» вообще нет слова эволюция, т.е. макроэволюция: Дарвин имел дело только с микроэволюцией, «происхождением видов». И с очевидным указанием на Дарвина первую свою крупную монографию Добржанский озаглавил «Генетика и происхождение видов»...
Василий Бабков. Путевые письма и микроэволюция Ф.Г. Добржанского. - Вавиловский журнал генетики и селекции (Вестник ВОГиС). М.: Институт цитологии и генетики СО РАН (Новосибирск), 2007. Том. 11. № 2. с.463—469
http://www.bionet.nsc.ru/vogis/pict_pdf/2007/t11_2/vogis_11_2_15.pdf

Форзац брошюры Т.Г. Моргана и Ю.А. Филипченко, из-во «Сеятель». Ленинград,1925. 58с.(Контакты Ю. А. Филипченко с Т. Х. Морганом вылились в публикацию небольшой брошюры «Наследственны ли приобретенные признаки»)
Ю.А. Филипченко (1882–1930) основал в 1919г. первую в нашей стране кафедру генетики. В 1912г., защитив магистерскую диссертацию как зоолог, он обратился к генетике. Уже в 1913г., 18 сентября доцент Филипченко приступил к чтению первого
в России курса генетики («Учение о наследственности и эволюция»), в 1915 г. издал первую свою книгу «Изменчивость и эволюция», в 1917г. — «Наследственность», в 1929г. на их основе вышел учебник «Генетика». В 1921г. он организовал при КЕПС Бюро по евгенике, превращенную позже в лабораторию генетики АН СССР. Его научные интересы охватывали: генетику качественных и количественных признаков, включая наследование таланта у человека, евгенику, генетические основы эволюции. Он предложил понятия микроэволюция и макроэволюция. Ю. А. Филипченко оставил обширную научную школу
Ю.А. Филипченко - ученый, педагог и организатор науки
https://cyberleninka.ru/article/n/yu-a-filipchenko-uchenyy-pedagog-i-organizator-nauki/viewer
1. Формирование исходного репертуара TCR на этапе рекомбинации — из какого сора...
Огромное разнообразие вариантов Т-клеточных рецепторов, как и разнообразие антител, формируется в результате относительно случайной «сборки» — событий рекомбинации, в ходе которых из имеющегося в геноме набора так называемых V-, D-, и J-сегментов выбирается по одному варианту.
Произвольная комбинация этих сегментов собирается в новый ген, а на стыке сегментов — в местах «сварки» — происходит дополнительное случайное удаление и добавление нуклеотидов. Таким образом, место стыка сегментов V и J (D-сегмент совсем короткий, и его фактически «заваривает» в середину) характеризуется наибольшей гетерогенностью. И, как правило, именно этот гипервариабельный участок в дальнейшем играет ключевую роль в распознавании антигена — является основной «бородкой» «ключа».
Однако после первичной сборки TCR большая часть (95–98%) Т-лимфоцитов погибает в тимусе в результате позитивной и негативной селекции, направленной на то, чтобы отобрать из множества случайно генерируемых последовательностей TCR работоспособные и безопасные для организма варианты. Эту жесткую селекцию пройдут только те TCR, которые, с одной стороны, принципиально способны распознавать антигены в контексте MHC, а с другой — не проявляют сильного взаимодействия с MHC, несущими собственные пептиды организма. Такой отбор необходим для того, чтобы избежать аутоиммунных реакций — атаки Т-клеток на собственные клетки хозяина.
Интересно, что существенная часть зрелых Т-лимфоцитов помимо генов функциональных TCR несет на второй (гомологичной) хромосоме также и последовательности нефункциональных цепей TCR со сбитой рамкой считывания либо стоп-кодоном, нарушающими синтез белка. Такие TCR не экспрессируются на поверхности клеток, а значит и не проходят позитивную и негативную селекцию в тимусе. Таким образом, эти бесполезные на первый взгляд последовательности могут нам многое рассказать о процессе рекомбинационных событий, в ходе которых рождается пре-селекционный репертуар TCR [13–17]. Сюда можно отнести и частоты используемых генных сегментов, из которых происходит не вполне случайная сборка TCR, и число случайно добавленных и «отгрызенных» нуклеотидов в местах соединения сегментов, и некоторые другие характеристики, определяющие исходную структуру репертуара TCR для каждого человека и в популяции.
На основе данных по репертуарам нефункциональных вариантов TCR была построена вероятностная модель сборки [16] и установлено нуклеотидное разнообразие уже для рабочих, функциональных вариантов пре-селекционного репертуара β-цепей TCR, составившее у людей (как популяции) порядка 10^13, что существенно ниже, чем предсказания более простых моделей, считающих процессы выбора сегментов и каждый случайно вставленный нуклеотид независимыми событиями [13] (рис. 3). Создание такой модели позволяет исследователям генерировать искусственные репертуары TCR на компьютере, практически точно копируя поведение реальной рекомбинационной машинерии человека.
Рисунок 3. Популяционное и индивидуальное разнообразие β-цепей TCR человека
Интересно, что пре-селекционный репертуар TCR однояйцевых (генетически идентичных) близнецов характеризуется практически идентичными частотами выбора генных сегментов [17], что указывает на решающее влияние индивидуальных генетических особенностей на статистику событий рекомбинации. В то же время, например, в парах мама/свой ребенок основные характеристики пре-селекционного репертуара TCR не ближе, чем таковые для пар мама/чужой ребенок [15]...
Дмитрий Чудаков. Анализ индивидуальных репертуаров Т-клеточных рецепторов. 11 дек. 2015
https://biomolecula.ru/articles/analiz-individualnykh-repertuarov-t-kletochnykh-retseptorov
Иммунология. К коровам. 7 белояра 2020
Атхарваведа VII, 79. К коровам
1 Богатые потомством, сверкающие на хорошем пастбище,
Пьющие чистую воду на хорошем водопое —
Пусть не овладеет вами ни вор, ни злоумышленник!
Пусть минует вас стрела Рудры!
2 Вы — знающие след, остающиеся (при нас),
Объединенные, носящие все имена.
Приходите ко мне, божественные, вместе с богами
В этот хлев, на это место!
Окропите вы нас жиром!
Атхарваведа. Избранное (Перевод, комментарии и вступ. статья Т.Я. Елизаренковой) 5.7Мб
http://www.bolesmir.ru/index.php?content=text&name=o285
1. Зарождение иммунологии
По происхождению иммунология — прикладная медицинская наука. Ее предыстория насчитывает более 2 тыс. лет. На протяжении этого времени основным подходом в данной области был эмпирический поиск путей предотвращения инфекционных заболеваний. В основе такого поиска было достоверное наблюдение о том, что люди, перенесшие некоторые «заразные болезни», повторно ими не заболевали. Ярко и очевидно этот факт проявлялся при оспе — именно это заболевание стало «плацдармом» для формирования иммунологии.
Описано предупреждение развития оспы у наследника китайского императора с помощью нанесения на слизистую носа материала из пустул людей, выздоравливающих от оспы. Это первое письменное свидетельство, которое однозначно можно отнести к области иммунологии (около 1 тыс. лет до н.э.). Профилактический опыт в данном случае был успешным. Подобный подход, позже названный европейцами вариоляцией (от лат. variola — оспа), был, по-видимому, независимо разработан в разных вариантах во многих регионах Азии. Во всяком случае, вариоляция была широко распространена к началу XVIII века, когда жена британского посла в Константинополе, леди Монтегю, в своих письмах (это был скорее роман в письмах) описала и пропагандировала эту процедуру. На протяжении XVIII века практика вариоляции, особенно после ее усовершенствования Р. Суттоном (R. Sutton), получила широкое распространение в Европе, вписавшись в идеологию эпохи Просвещения. Европейские монархи подвергали вариоляции своих детей и внуков в знак причастности прогрессивным веяниям времени (по настоянию Екатерины II оспа была привита ей и ее сыну — будущему императору Павлу I). Надо признать, что вариоляция несла значительный риск и нередко приводила к развитию заболевания и смерти, что в конце концов было осознано европейцами, не готовыми жертвовать даже единичными
гражданами ради абстрактного прогресса (в азиатской традиции с большей легкостью относятся к индивидуальным утратам). Вариоляция была запрещена в большинстве стран.
В 90-х годах XVIII века английский врач Э. Дженнер (E. Jenner), работавший в сельской скотоводческой местности, сделал наблюдение, что коровницы, контактировавшие со скотом, больным коровьей оспой, если и заболевали человеческой оспой, то переносили ее легко (самим коровницам этот факт был известен давно). Основываясь на этом подтвержденном, но все-таки эмпирическом факте, Э. Дженнер предпринял рискованный эксперимент на человеке: он привил коровью оспу от заболевшей ею коровницы 8-летнему Джеймсу Фиппсу. Реакция на прививку проявилась в виде кратковременного недомогания. Повторная прививка коровьей оспы дала еще более слабую реакцию. После этого Дженнер предпринял шаг, который в настоящее время мог бы быть расценен как преступление: привил подростку материал от больного человеческой оспой. Заболевание не развилось. Отчет об этом успешном опыте вакцинации (от латинского vacca — корова) был опубликован в научной прессе в 1796г. Однако эту публикацию нельзя расценивать как начало иммунологии, поскольку речь в ней шла о конкретной профилактической процедуре, а не об общих принципах и правилах, которые можно было бы расценивать как фундамент новой науки.
На протяжении XIX века вакцинация приобрела широкое распространение в цивилизованном мире и применялась в практически неизменном виде до недавнего времени, когда международное сообщество признало факт элиминации оспы (1980)...
Данная книга первоначально задумывалась как переработанный и обновленный вариант учебника «Основы иммунологии», вышедшего более десяти лет тому назад (в 1999 г.) в издательстве «Медицина». Однако изменения, которые произошли за этот срок в иммунологии, столь глубоки, что потребовали не просто обновления или дополнения старого текста новыми сведениями, а создания новой книги. В этом читатель сможет убедиться, сопоставляя структуры старой и новой книг или, проще, сравнивая их оглавления...
А.А. Ярилин. Иммунология. 2010. 749с.
https://vk.com/doc399489626_539293729
История научной иммунологии насчитывает чуть более 100 лет. Несмотря на молодость, она относится к наиболее значимым и «влиятельным» наукам медико-биологического профиля. Ей принадлежит одно из первых мест по числу профильных журналов, издаваемых в мире, и объему помещаемых в них публикаций. Во всех странах мира иммунология вошла в число самостоятельных вузовских дисциплин относительно недавно, лишь в 70-е, а в широком масштабе — в 80-е годы. Мне сопутствовала удача, и в 1971 г. удалось организовать первую в нашей стране кафедру иммунологии во II Московском медицинском институте им. Н.И. Пирогова.
А.А. Ярилин. Основы иммунологии. Учебник. 1999. 608с. djvu
https://vk.com/doc399489626_539276779
Взаимосвязь врожденного и адаптивного иммунитета. К Соме-Рудре
РигВеда VI, 74. К Соме-Рудре
1 О Сома-Рудра, сохраняйте (вашу) асурскую силу,
Пусть наши желания вовремя вас достигнут!
В каждый дом принося семь сокровищ,
Будьте на благо (нашему) двуногому, (нашему) четвероногому!
2 О Сома-Рудра, вырвите (и рассейте) в разные стороны
(Ту) болезнь, которая проникла в наш дом!
Прогоните далеко прочь Гибель!
Да будут у нас (вещи,) приносящие счастье (и) добрую славу!
3 О Сома-Рудра, даруйте нам все эти
Целебные средства, (даруйте их нашим) телам!
Развяжите, освободите нас от содеянного греха,
Который привязан к нашим телам!
4 Вы двое с острым оружием, с острым дротиком, очень милые,
О Сома-Рудра, очень нас пожалейте здесь!
Освободите нас от петли Варуны!
Охраняйте нас благожелательно!
Ригведа. Мандалы V-VIII (перевод Т.Я. Елизаренковой) 8.7Мб
https://vk.com/doc425763081_532185721

Рис. 1.8. Взаимосвязь врожденного и адаптивного иммунитета. Врожденный иммунитет обеспечивает презентацию антигена и костимуляцию, необходимые для запуска адаптивного иммунитета. В свою очередь, адаптивный иммунитет благодаря выработке антител и цитокинов придает реакциям врожденного иммунитета избирательность действия и повышает их эффективность (с.44)
...уникальная особенность иммунологии состоит в том, что ее «сердцевина» не вполне стабилизировалась и меняется во времени в большей степени, чем основы других наук. Достаточно сказать, что на протяжении полувека парадигма иммунологии менялась по меньшей мере два раза — сначала при рождении «неинфекционной» (по преимуществу клеточной) иммунологии в 50–60-е годы ХХ века и затем — совсем недавно, при формировании новых представлений об иерархии и взаимодействии врожденного и адаптивного иммунитета (с.11)
...Наиболее крупным теоретическим обобщеним, повлекшим большое число экспериментальных исследований и практически значимых разработок, послужило учение Ч. Джанеуея (Ch. Janeway) и его последователей о природе распознавания во врожденном иммунитете и иерархических взаимодействиях врожденного и адаптивного иммунитета. При этом, во-первых, был открыт новый тип иммунологического распознавания, заставивший отказаться от представлениий о неспецифичности врожденного иммунитета, во-вторых, было обосновано представление о невозможности запуска адаптивного иммунитета без предварительной активации врожденного иммунитета. Исследования, проводимые в области иммунологии в ХХI веке, в большей или меньшей степени ориентированы на эту концепцию (с.23)
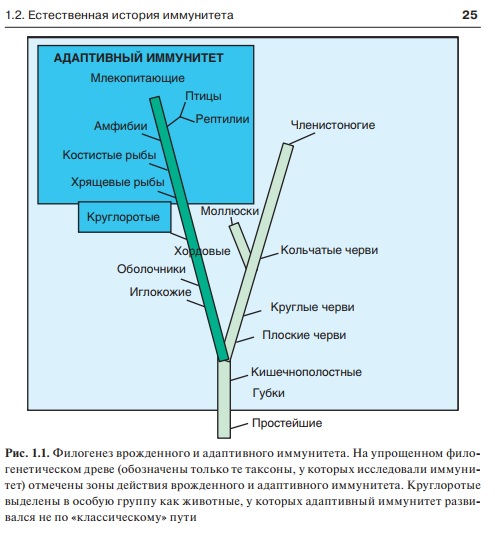
Рис. 1.1. Филогенез врожденного и адаптивного иммунитета. На упрощенном филогенетическом древе (обозначены только те таксоны, у которых исследовали иммунитет) отмечены зоны действия врожденного и адаптивного иммунитета. Круглоротые выделены в особую группу как животные, у которых адаптивный иммунитет развивался не по «классическому» пути
Так, схематично можно представить систему иммунитета, которую принято называть врожденной. Эта форма иммунитета характерна для всех многоклеточных животных (в несколько иной форме — и для растений). Ее возраст — 1,5 млрд лет. Система врожденного иммунитета весьма эффективно защищала первичноротых многоклеточных животных, а также низших вторичноротых, часто имевших крупные размеры (рис. 1.1). Проявления врожденного иммунитета на разных стадиях эволюции и в разных таксонах чрезвычайно разнообразны. Однако общие принципы его функционирования одинаковы на всех стадиях развития многоклеточных. Главные составляющие врожденного иммунитета:
распознавание чужеродных агентов во внутренней среде организма с помощью рецепторов, специализированных на узнавании «образов» патогенности;
элиминация опознанных чужеродных агентов из организма путем фагоцитоза и расщепления.
У хордовых произошло скачкообразное формирование другой разновидности иммунитета: примерно 500 млн лет назад возник адаптивный (т.е. приспособительный) или приобретенный иммунитет. Ветвь адаптивного иммунитета, получившая интенсивное развитие, зародилась у хрящевых рыб. Особый вариант адаптивного иммунитета, основанный на использовании других распознающих и эффекторных молекул, обнаружен у более примитивных хордовых — круглоротых. Адаптивный иммунитет тесно связан с врожденным и во многом основывается на его проявлениях. Однако эти типы иммунитета сильно различаются (табл. 1.2).


Существенное отличие адаптивного иммунитета от врожденного — способ распознавания чужого (табл. 1.3). В адаптивном иммунитете оно осуществляется при помощи молекул особого типа (иммуноглобулинов или других белков суперсемейства иммуноглобулинов), при этом распознаются не паттерны, а индивидуальные молекулы или небольшие группы сходных молекул, называемые антигенами. Существует порядка 10^6 различных антигенов. Такое число рецепторов не только не может быть представлено на одной клетке, но и не может быть закодировано в геноме позвоночных, содержащем только десятки тысяч генов. Именно поэтому в процессе эволюции адаптивного иммунитета сформировался сложный механизм генерации разнообразия антигенспецифических рецепторов: при развитии специализированных клеток (лимфоцитов), происходит перестройка их генов, кодирующих антигенраспознающие рецепторы, что приводит к образованию в каждой клетке рецептора с уникальной специфичностью. При активации каждая клетка может дать начало клону, все клетки которого будут иметь рецепторы той же специфичности. Таким образом, каждый конкретный антиген распознают не все лимфоциты, а только отдельные их клоны, имеющие специфические антигенраспознающие рецепторы.
Если паттерн-распознающие рецепторы врожденного иммунитета образовались в процессе эволюции как молекулы, распознающие чужеродные, но не собственные молекулы организма, то специфичность антиген-распознающих рецепторов системы адаптивного иммунитета формируется случайно. Это потребовало развития дополнительных механизмов селекции для устранения «ненужных» и «опасных» (направленных против «своего»)клонов лимфоцитов. Такие механизмы достаточно эффективны, однако все же не полностью устраняют риск развития аутоиммунных процессов — иммунных реакций, направленных против собственных антигенов, вызывающих повреждение организма хозяина.
Оба типа иммунитета образуют целостную систему, при этом врожденный иммунитет служит фундаментом для развития адаптивного. Так, лимфоциты распознают антиген в процессе презентации, осуществляемой преимущественно клетками врожденного иммунитета. Удаление из организма антигена и несущих его клеток происходит с помощью реакций, в основе которых лежат механизмы врожденного иммунитета, получившие специф






