|
Повышенное артериальное д.
| | | ИФР - 1
| белок из семейства инсулиноподобных факторов роста. ИФР-1 состоит из одной полипептидной цепи длиной 70 аминокислотных остатков с тремя внутримолекулярными дисульфидными мостиками.
| производится гепатоцитами печени в ответ на стимуляцию их соматотропиновых рецепторов
|
| ИФР-1 также обеспечивает обратную связь с гипоталамусом и гипофизом по соматотропной оси: от уровня ИФР-1 в крови зависит секреция соматотропин-рилизинг-гормона и соматотропного гормона. При низком уровне ИФР-1 в крови секреция соматотропин-рилизинг-гормона и соматотропина возрастает, при высоком — снижается. Также ИФР-1 регулирует секрецию соматостатина: высокий уровень ИФР-1 приводит к возрастанию секреции соматостатина, низкий — к её снижению. Этот механизм является ещё одним способом регуляции уровня соматотропного гормона в крови. Но действие может быть заторможено недостаточным питанием, нечувствительностью гормона роста, отсутствием реакции рецепторов, или неудачным, ниже необходимого минимума сигнального пути, сообщением рецепторов. В экспериментах на крысах было обнаружено, что количество ИФР-1 и РНК в печени положительно связано с недостатком казеина и отрицательно связано с недостатком белка в пище. Также было установлено, что при недостатке ИФР-1 в крови он может продуцироваться в самих мышцах.
Уровень ИФР-1 в крови зависит от действия на печень не только соматотропного гормона, но и половых стероидов и тиреоидных гормонов, глюкокортикоидов, инсулина. При этом инсулин, андрогены, эстрогены повышают секрецию ИФР-1 печенью, а глюкокортикоиды её снижают. Это является одной из причин синергизма инсулина, соматотропина, половых и тиреоидных гормонов в отношении процессов роста и развития организма, роста и дифференцировки тканей, и одной из причин характерного тормозящего действия глюкокортикоидов на процессы линейного роста, полового созревания и пр. ИФР-1 воздействует на развитие всю жизнь, но его уровень в крови не постоянный: наиболее низкий уровень ИФР-1 производства в детстве и в старости, а самый высокий — во время подросткового периода жизни.
|
ГР/ИФР1 каскад сильно консервативен у различных групп животных (позвоночных и беспозвоночных). В общих чертах он заключается в том, что ИФР1 через свой рецептор запускает каскад реакций, приводящих к ингибированию факторов транскрипции (DAF-16 у C.elegans или FOXO1 у мышей). Вероятно, эти факторы транскрипции регулируют экспрессию генов, которые увеличивают продолжительность жизни
Если у беспозвоночных животных insulin/ИФР1 — путь один, у высших позвоночных, в том числе, млекопитающих, этот путь подразделяется на два. Эти два пути имеют перекрывающиеся функции, но инсулин главным образом участвует в регуляции метаболизма, а ГР/ИФР1 путь играет важную роль в процессах роста, развития и, возможно длительности жизни.
Именно гены ИФР-каскада стали первыми открытыми «генами старения» — то есть генами, повреждение которых привело к увеличению продолжительности жизни
|
Старение
|
Рак
| |
| Лейкотриены
| эйкозаноид 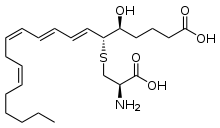
| 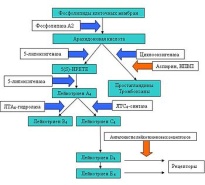 лейкотриены образуются из арахидоновой кислоты, которая, в свою очередь, отщепляется от фосфолипидов цитоплазматической мембраны, с помощью фермента фосфолипаза A2.
Далее арахидоновая кислота может трансформироваться двумя путями: под влиянием ЦОГ она превращается в простаноиды, а под влиянием липоксигеназной ферментной системы в лейкотриены.
Липоксигеназная ферментная система относится к растворимым цитозольным ферментам, они обнаружены в цитоплазме альвеолярных макрофагах, тромбоцитах, тучных клетках и лейкоцитах. Наиболее важным среди ферментов этой системы является 5-липоксигеназа (5-ЛОГ). Активация перечисленных клеток приводит к перемещению 5-ЛОГ к мембране ядерного аппарата и связыванию со специфическим белком — 5-ЛОГ-активирующим протеином (5-ЛОГ-АП). 5-ЛОГ-АП является кофактором при взаимодействии арахидоновой кислоты и 5-ЛОГ. Таким образом, арахидоновая кислота под воздействием комплекса 5-ЛОГ + 5-ЛОГ-АП превращается в нестабильное соединение 5-гидроксипероксиэйкозатетраеновую кислоту (5-HPETE), из которой в свою очередь образуется ЛТА4. Обе эти реакции катализируются активированной 5-ЛОГ, расположенной на перинуклеарной мембране.
Далее ЛТА4 может превращаться двумя путями: либо при участии цитозольного фермента ЛТА4-гидролазы в ЛТВ4, либо под воздействием ЛТС4-синтетазы с образованием цисЛТС4. ЛТС4 выходит во внеклеточное пространство и далее с помощью g-глутамилтрансептидазы превращается в ЛТD4, который затем под влиянием дипептидазы образует ЛТЕ4. ЛТЕ4 является субстратом для образования ЛТF4.
лейкотриены образуются из арахидоновой кислоты, которая, в свою очередь, отщепляется от фосфолипидов цитоплазматической мембраны, с помощью фермента фосфолипаза A2.
Далее арахидоновая кислота может трансформироваться двумя путями: под влиянием ЦОГ она превращается в простаноиды, а под влиянием липоксигеназной ферментной системы в лейкотриены.
Липоксигеназная ферментная система относится к растворимым цитозольным ферментам, они обнаружены в цитоплазме альвеолярных макрофагах, тромбоцитах, тучных клетках и лейкоцитах. Наиболее важным среди ферментов этой системы является 5-липоксигеназа (5-ЛОГ). Активация перечисленных клеток приводит к перемещению 5-ЛОГ к мембране ядерного аппарата и связыванию со специфическим белком — 5-ЛОГ-активирующим протеином (5-ЛОГ-АП). 5-ЛОГ-АП является кофактором при взаимодействии арахидоновой кислоты и 5-ЛОГ. Таким образом, арахидоновая кислота под воздействием комплекса 5-ЛОГ + 5-ЛОГ-АП превращается в нестабильное соединение 5-гидроксипероксиэйкозатетраеновую кислоту (5-HPETE), из которой в свою очередь образуется ЛТА4. Обе эти реакции катализируются активированной 5-ЛОГ, расположенной на перинуклеарной мембране.
Далее ЛТА4 может превращаться двумя путями: либо при участии цитозольного фермента ЛТА4-гидролазы в ЛТВ4, либо под воздействием ЛТС4-синтетазы с образованием цисЛТС4. ЛТС4 выходит во внеклеточное пространство и далее с помощью g-глутамилтрансептидазы превращается в ЛТD4, который затем под влиянием дипептидазы образует ЛТЕ4. ЛТЕ4 является субстратом для образования ЛТF4.
| «Пептидные лейкотриены» модулируют специфические рецепторы, сопряженные с G-белком. Их обозначают CysLT-R. В настоящее время выделяют 2 типа CysLT. Взаимодействие лейкотриенов с рецепторами 1 типа (CysLT1) определяет спектр их основных эффектов (бронхоспазм). Связывание ЛТ с рецепторами 2 типа (CysLT2) изменяет тонус и проницаемость сосудов.
Лейкотриен B4 модулирует другой тип рецепторов — BLT1- и BLT2-рецепторы (другое название LTB4-рецепторы).
| - ЛТB4 — опосредует хемотаксис, экссудацию плазмы, сокращение паренхимы лёгких, участие в иммунных ответах.
- ЛТC4, ЛТD4, ЛТE4 являются основными компонентами МРСА, поэтому в первую очередь эта группа лейкотриенов относится к мощным бронхоконстрикторам. Также эти лейкотриены способны повышать тонус гладких мышц ЖКТ, опосредовать экссудацию плазмы и сокращение паренхимы лёгких.
Роль в патологии
- Лейкотриены участвуют в патогенезе бронхиальной астмы. Вместе с гистамином лейкотриены относятся к медиаторам ранней фазы аллергической реакции немедленного типа. В результате действия гистамина возникает мгновенный и кратковременный бронхоспазм, лейкотриены же вызывают отсроченный и более длительный бронхоспазм.
- Лейкотриены вызывают очень неприятный аспириновый бронхоспазм, возникающий при приёме неселективных НПВС: аспирин и др.[3] Аспириновый бронхоспазм порождается следующим образом: ЦОГ, которую ингибируют НПВС, катализирует реакцию превращения арахидоновой кислоты в циклический эндопероксид ПГ H2. Это ведёт к тому, что синтез ПГ резко уменьшается, а на этом фоне преобладающими станут лейкотриены. Однако, активность фосфолипазы A2 остаётся неизменной, соответственно арахидоновой кислоты отщепляется от фосфолипидов цитоплазматической мембраны столько же, сколько и в норме. Если в физиологических условиях арахидоновая кислота равномерно распределялась, поступая на синтез и ПГ и ЛТ, то при ингибировании ЦОГ, она целиком и полностью будет поступать на синтез лейкотриенов. Таким образом, при использовании неселективных НПВС, в тканях человека будет не просто вакатное (лат. vacuus — пустой) преобладание ЛТ, но они будут синтезироваться интенсивнее, чем в норме. ЛТC4, ЛТD4, ЛТE4 входят в комплекс МРСА, которая порождает бронхоспазм.
|
|
|
Астма, удушье
| |
| Тромбоксаны
| 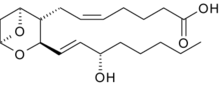 Тромбоксан A2
Тромбоксан A2
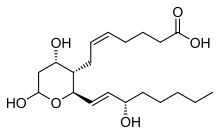 Тромбоксан B2
Тромбоксан B2
| Синтез происходит в тромбоцитах под влиянием фермента тромбоксан-А синтазы из эндоперекисей, образующихся из арахидоновой кислоты с помощью фермента циклооксигеназы. Период полураспада тромбоксана А2 в организме человека составляет 30 секунд.
| Тромбоксаны взаимодействуют с рецепторами, сопряженными с G белком (а именно с Gq белком).
| Тромбоксаны сужают сосуды, повышают артериальное давление и активируют агрегацию тромбоцитов. Концентрация тромбоксанов находится в равновесии с уровнем их антагониста простациклина.
| Тромбоксан А2 синтезируется активированными тромбоцитами. Стимулирует активацию новых тромбоцитов и их агрегацию. Агрегация тромбоцитов достигается повышением уровня экспрессии гликопротеинового комплекса GP IIb/IIIa на их мембранах. Циркулирующий фибриноген связывается с этим комплексом, укрепляя тромб.
|
| Считается, что сужение коронарных сосудов при стенокардии Принцметала вызвана тромбоксанами.
| |
|
Гормоны ЖКТ
| |
| Соматостатин
|
| Желудок, проксимальный отдел тонкой кишки, поджелудочная железа D-клетки
|
| Тормозит выделение инсулина и глюкагона, большинства известных желудочно-кишечных гормонов (секретина, ГИПа, мотилина, гастрина); тормозит активность париетальных клеток желудка и ацинарных клеток поджелудочной железы
|
|
|
| |
| Вазоактивный интестинальный (ВИП) пептид
|
| Во всех отделах желудочно-кишечного тракта D-клетки
|
| Тормозит действие холецистокинина, секрецию соляной кислоты и пепсина желудком, стимулированную гистамином, расслабляет гладкие мышцы кровеносных сосудов, желчного пузыря
|
|
|
| |
| Панкреатический полипептид (ПП)
|
| Поджелудочная железа D2-клетки
|
| Антагонист ХЦК-ПЗ, усиливает пролиферацию слизистой оболочки тонкой кишки, поджелудочной железы и печени; участвует в регуляции обмена углеводов и липидов
|
|
|
| |
| Гастрин
|
| Антральная часть желудка, поджелудочная железа, проксимальный отдел тонкой кишки G-клетки
|
| Стимулирует секрецию и выделение пепсина желудочными железами, возбуждает моторику расслабленного желудка и двенадцатиперстной кишки, а также желчного пузыря
| Секреция гастрина повышается в ответ на холинергическую (блуждающим нервом) и в меньшей степени на симпатическую стимуляцию желудка. Также секреция гастрина повышается инсулином, гистамином, присутствием в желудке или в плазме крови олигопептидов и свободных аминокислот — продуктов расщепления белков. Увеличение секреции гастрина в ответ на олигопептиды и свободные аминокислоты, а также в ответ на симпатическую или холинергическую стимуляцию является физиологическим механизмом инициации пищеварения при виде и запахе пищи или при поступлении пищи в желудок. Секреция гастрина также повышается при гиперкальциемии.
Угнетается секреция гастрина высоким уровнем соляной кислоты в желудке (что является одной из отрицательных обратных связей, регулирующих секрецию гастрина), простагландином Е, эндогенными опиоидами — эндорфинами и энкефалинами, аденозином, кальцитонином. Сильно угнетает секрецию гастрина соматостатин, одновременно угнетающий секрецию других панкреатических и кишечных пептидов — холецистокинина, секретина, ВИП и др. Повышение гастрином секреции соматостатина, угнетающего секрецию гастрина, является ещё одним примером отрицательной обратной связи.
Секреция гастрина также угнетается холецистокинином и секретином. Физиологическое значение этого механизма состоит в уменьшении секреции кислоты и пепсина после начала кишечной фазы пищеварения и обеспечении функционального покоя опустошившегося желудка, а также в обратной связи, ограничивающей гиперсекрецию кислоты (поскольку уровни секретина и холецистокинина зависят от pH поступающей в двенадцатиперстную кишку пищевой кашицы).
|
| гипертрофия слизистой желудка, усиление её складчатости, функциональную гиперплазию желёз желудка
| |
| Гастрон
|
| Антральный отдел желудка G- клетки
|
| Снижает объем желудочной секреции и выход кислоты в желудочном соке
|
|
|
| |
| Бульбогастрон
|
| Антральный отдел желудка G- клетки
|
| Тормозит секрецию и моторику желудка
|
|
|
| |
| Дуокринин
|
| Антральный отдел желудка G- клетки
|
| Стимулирует выделение секрета бруннеровых желез двенадцатиперстной кишки
|
|
|
| |
| Секретин
| пептидный гормон
| Тонкий кишечник S-клетки
| поджелудочная железа
| Стимулирует секрецию бикарбонатов и воды поджелудочной железой, печенью, железами Бруннера, пепсина; тормозит секрецию в желудке
|
|
|
| |
| Холецистокинин-панкреозимин (ХЦК-ПЗ)
|
| Тонкий кишечник I-клетки
|
| Возбуждает выход ферментов и в слабой степени стимулирует выход бикарбонатов поджелудочной железой, тормозит секрецию соляной кислоты в желудке, усиливает сокращение желчного пузыря и желчевыделение, усиливает моторику тонкой кишки
|
|
|
| |
| Бомбезин (гастринвысвобождающий пептид)
|
| Желудок и проксимальный отдел тонкой кишки Р-клетки
|
| Стимулирует высвобождение гастрина, усиливает сокращение желчного пузыря и выделение ферментов поджелудочной железой, усиливает выделение энтероглюкагона
|
|
|
| |
| Энтероглюкагон
|
| Тонкий кишечник ЕС1-клетки
|
| Тормозит секреторную активность желудка, снижает в желудочном соке содержание К+ и повышает содержание Са2+, тормозит моторику желудка и тонкой кишки
|
|
|
| |
| мотилин
|
| Проксимальный отдел тонкой кишки ЕС2-клетки
|
| Возбуждает секрецию пепсина желудком и секрецию поджелудочной железы, ускоряет эвакуацию содержимого желудка
|
|
|
| |
| Гастроингибирующий пептид (ГИП)
|
| Тонкий кишечник К-клетки
|
| Тормозит выделение соляной кислоты и пепсина, высвобождение гастрина, моторику желудка, возбуждает секрецию толстой кишки
|
|
|
| |
| Нейротензин
|
| Дистальный отдел тонкой кишки N-клетки
|
| Тормозит секрецию соляной кислоты железами желудка, усиливает высвобождение глюкагона
|
|
|
| |
| Энкефалины (эндорфины)
|
| Проксимальный отдел тонкой кишки и поджелудочная железа L-клетки
|
| Тормозит секрецию ферментов поджелудочной железой, усиливает высвобождение гастрина, возбуждает моторику желудка
|
|
|
| |
| Субстанция Р
|
| Тонкая кишка ЕС1-клетки
|
| Усиливает моторику кишечника, слюноотделение, тормозит высвобождение инсулина
|
|
|
| |
| Вилликинин
|
| Двенадцатиперстная кишка ЕС1-клетки
|
| Стимулирует ритмические сокращения ворсинок тонкой кишки
|
|
|
| |
| Энтерогастрон
|
| Двенадцатиперстная кишка ЕС1-клетки
|
| Тормозит секреторную активность и моторику желудка
|
|
|
| |
| | | | | | | | | | | | | | | |






