

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...

Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...
Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого...
Состав сооружений: решетки и песколовки: Решетки – это первое устройство в схеме очистных сооружений. Они представляют...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Отражение на счетах бухгалтерского учета процесса приобретения: Процесс заготовления представляет систему экономических событий, включающих приобретение организацией у поставщиков сырья...
Эволюция кровеносной системы позвоночных животных: Биологическая эволюция – необратимый процесс исторического развития живой природы...
Интересное:
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Финансовый рынок и его значение в управлении денежными потоками на современном этапе: любому предприятию для расширения производства и увеличения прибыли нужны...
Национальное богатство страны и его составляющие: для оценки элементов национального богатства используются...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
2.1 Молекулярная характеристика генов FLO (львиный зев) и LFI (арабидопсис)
Ген львиного зева FLO был клонирован одним из первых генов, влияющих на развитие цветка. Для его клонирования использовали мутант По-613, образующий генеративные соцветия вместо цветков. Однако гомозиготы по flo-613 изредка формировали нормальные цветки. Из семян этих цветков развивались растения дикого типа. Значит, мутация flo-613 генетически нестабильна и иногда ревертировала к аллели дикого типа. Реверсия коррелировала с эксцизией транспозона ТатЗ и, следовательно, мутантный фенотип был обусловлен инсерцией этого мобильного элемента в ген FLO. Таким образом, ген FLO был клонирован с использованием последовательности транспозона ТатЗ в качестве пробы. У всех ревертантов к дикому типу при использовании клонированного фрагмента гена FLO как пробы для гибридизации появлялся фрагмент длиной 4т. п. о. Такой же длиной обладал гибридизовавшийся с FLO фрагмент исходного растения дикого типа, использованного для транспозонового мутагенеза. При этом у некоторых ревертантов сохранялся фрагмент длиной 7.5 т. п. о., такой же, как и у мутанта flo-613. Вероятно, среди ревертантов были как гетерозиготные, так и гомозиготные растения.
Однако при гибридизации ДНК мутантных растений среди них тоже были выявлены гетерозиготы, у которых присутствовали оба фрагмента. Так, при секвенировании сайта эксцизии ТатЗ из гетерозиготы с фенотипом По обнаружили инсерцию 8 пар оснований. Таким образом, неточная эксцизия ТатЗ приводила к изменению рамки считывания, и мутантный фенотип сохранялся.
В сайте эксцизии у ревертанта отмечены замены двух пар оснований, и не обнаружены инсерции или делеции нуклеотидов. В потомстве от самоопыления всех гетерозиготных ревертантов имеются растения дикого типа и мутанты в соотношении 3:3. Потомство большинства гомозиготных ревертантов состояло только из растений дикого типа.
|
|
Мутация flo-613, как и многие другие гомеозисные мутации львиного зева, использованные для клонирования генов развития цветка и о которых пойдет речь в этой главе, была получена в массивном эксперименте по транспозоновому мутагенезу. Растения львиного зева с высокоактивными мобильными элементами Тат (от Transposone of Antirrhinum majus) выращивали при температуре 15 °С. Именно при этой температуре для мобильных элементов группы Тат наблюдается наибольшая частота транспозиций. Многие из 26000 потомков Ml этих растений, полученных при самоопылении, содержали рецессивные мутации в гетерозиготном состоянии. Такие рецессивные мутации можно было выявить в следующем поколении М2 после самоопыления растений Ml. И в самом деле, среди 80000 растений М2 было обнаружено более 15 независимых гомеозисных мутаций, затрагивающих развитие цветка (рис.2).
Ген арабидопсиса LFY был клонирован по гомологии с геном львиного зева FLO. В отличие от многих других гомеозисных генов, затрагивающих развитие цветка, FLO и LFY, похоже, уникальные гены и не являются членами генного семейства. Кодируемые ими белки были на 70 % гомологичны друг другу и не обладали значительной гомологией ни с одним из известных на момент их клонирования белков. Наличие богатого пролином домена на N-конце и кислой области в центральной части белка свидетельствуют о том, что, вероятно, они могут служить активаторами транскрипции. Подтверждается это и тем, что белок Lfy локализован в основном в ядре. Характер их экспрессии очень похож. РНК FLO и LFY начинает накапливаться в закладывающихся зачатках цветка на границе цветочной меристемы и не обнаруживается в генеративной меристеме. В момент приобретения цветочными меристемами морфологических различий (стадии 1 и 2) экспрессия усиливается в равной степени по всему молодому зачатку цветка. На стадиях 3 и 4, когда индуцируется экспрессия генов идентичности органов, РНК FLO и LFY уже практически не обнаруживается в центре начинающего формироваться цветка. Из известных генов они первыми начинают экспрессироваться в клетках, из которых затем сформируется цветок.
|
|
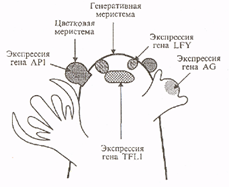
Рис.2. Схематическое изображение генеративного побега арабидопсиса и начальные этапы экспрессии генов, регулирующие развитие цветка.
Однако в характере экспрессии FLO и LFY есть одно важное отличие, помогающее понять, как развиваются меристемы. Различие является следствием того, что у цветка львиного зева есть прицветники, а у цветка арабидопсиса их нет. Ген FLO экспрессиру-ется как в примордиях прицветников, так и в цветковых меристемах. В то же время для гена LFY не наблюдается экспрессия в клетках, окружающих цветковые меристемы, и, вероятно, соответствующих клеткам львиного зева, из которых образуются прицветники. У мутантов Ify на месте цветков дикого типа без прицветников образуются генеративные побеги или «цветки» с околоплодными листьями, которые можно отождествлять с прицветниками. Значит, продукт LFY дикого типа подавляет инициацию или образование прицветников. Предполагают, что ген LFY влияет на судьбу группы клеток на периферии цветочной меристемы: в цветках дикого типа все или большинство этих клеток формируют цветочную меристему, тогда как у мутантов Ify эти клетки дают начало меристеме и примордию прицветника. В этом отношении мутант Ify похож на дикий тип растений таких видов, как Antirrhinum majus, у которых из одного зачатка образуются меристема цветка и прицветник. Считается, что наличие прицветников - эволюционно более древний признак.
Насколько универсальны охарактеризованные механизмы индукции цветка? Пока что на этот вопрос ответить не просто. Похоже, что для табака, который является более близким родственником львиного зева, чем арабидопсис, индукция развития цветковой меристемы происходит по-другому. Гены табака Nicotiana tabacum NFL1 и NFL2, гомологи генов FLO и LFY, транскрибируются во время как вегетативного, так и репродуктивного развития растений. Их экспрессия не инициируется во время индукции цветка, как это характерно для генов FLO и LFY. Возможно, это связано с тем, что табак имеет терминальный цветок, в то время как соцветия львиного зева и арабидопсиса не детерминированы. Трансгенные растения арабидопсиса, в которых конститутивно экспрессируется ген LFY, тоже имеют терминальный цветок. В геномах диплоидных растений львиного зева и арабидопсиса имеется только одна копия гена FLO или LFY. Табак же является аллотет-раплоидом, и каждый из его диплоидных геномов содержит по одной копии гомологичного FLO/LFY гена.
|
|
2.2 Молекулярная характеристика генов SQUA (львиный зев) и AP 1 (арабидопсис)
Ген львиного зева SQUA был клонирован с использованием в качестве пробы гена DEF из этого же растения, содержащего MADS-бокс (MADS-бокс — это последовательность ДНК, кодирующая домен MADS. Он получил свое название от четырех клонированных первыми генов с этим боксом: МСМ1, AGAMOUS, DEFICIENS, SRF. Домен MADS представляет собой консервативную область белков - факторов транскрипции, участвующую в связывании с ДНК (подробнее об этом см. далее). По гомологии с DEF было выделено 9 независимых генов львиного зева. Один из клонов при гибридизации геномных блотов по Саузерну проявлял рестрикционный полиморфизм для ДНК дикого типа и нестабильной мутации squa-29. Ревертанты SQUA имели фрагмент ДНК такой же длины, как и растения дикого типа.
Ген арабидопсиса API был клонирован как новый член семейства генов, содержащих MADS-бокс.
Белки Squa и Apl характеризуются 68%-ной гомологией. Наличие в этих белках домена MADS предполагает, что они могут специфически связываться с ДНК и функционировать как активаторы транскрипции. РНК SQUA и API индуцируется в молодых цветочных примордиях, как только они становятся видимыми на границе генеративной меристемы (чуть позже, чем РНК FLO и LFY). На стадиях I и 2 РНК SQUA и API однородно экспрессируется по всему примордию цветка (рис. 11.33), на стадии 3 экспрессия снижается в двух внутренних мутовках, и затем РНК API обнаруживается только в примордиях чашелистиков и лепестков. РНК SQUA, кроме того, менее интенсивно экспрессируется и в примордиях прицветников и карпел. В вегетативных органах, за исключением самых верхних листьев, примыкающих к соцветию, РНК SQUA не обнаруживается (рис. 11.32, в).
|
|
Ген SQUA экспрессируется в мутантах flo, а ген FLO — в мутантах squa. Таким образом, FLO и SQUA активируются независимо друг от друга. Подобным образом, ген LFY экспрессируется в мутантах apl. Фенотип двойных мутантов flo squa и Ify apl выражен значительно сильнее, чем фенотип одиночных мутантов по этим генам, что также свидетельствует о независимом и синергид-ном действии этих генов в процессе формирования цветковой меристемы и о частичной вырожденности их активностей.
Ген LFY не только необходим для формирования цветка, но и достаточен для определения идентичности цветка, т.е. развития меристемы как цветковой меристемы. Основной побег трансгенных растений p35S::LFY с конститутивной экспрессией этого гена переходит к цветению раньше, чем растения дикого типа, а вторичные побеги превращаются в цветки. Хотя ген API экспрессируется несколько позднее, чем LFY, он тоже необходим для инициации развития цветка. Конститутивная экспрессия трансгена p35S::APl тоже ускоряет переход растения к цветению и приводит к образованию цветков на месте вторичных побегов. По-видимому, гены LFY/AP1 и FLO/SQUA оказывают наибольшее влияние на инициацию цветковых меристем. Однако в этот процесс вносят определенный вклад и другие гены.
|
|
|

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Адаптации растений и животных к жизни в горах: Большое значение для жизни организмов в горах имеют степень расчленения, крутизна и экспозиционные различия склонов...

Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!