Из схемы видно их сходство. Все они зависят от образования связанных с клеточными мембранами комплексов фермент-кофактор.
Ломанная кривая обозначает участки факторов VII, IX, X и протромбина, содержащие гамма-карбоксипгутаминовую кислоту и связывающие Са2+ и фосфолипиды.
Штриховкой обозначены белки, связывающиеся с мембранами с помощью гидрофобного взаимодействия.
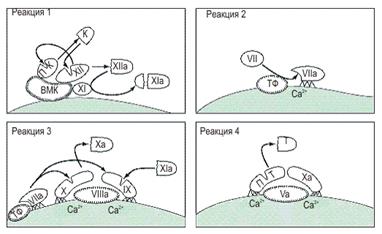
Схема основных реакций свертывания крови
ВМК - высокомолекулярный кининоген;
К - калликреин;
ПК - прекалликреин;
ПТ - протромбин;
Т - тромбин;
ТФ - тканевой фактор.
Каждая из этих реакций представляет собой образование активной протеазы из ее предшественника путем протеолиза; все они идут на фосфолипидных мембранах, требуют присутствия Са2+ и кофакторов.
В реакции 1 (внутренний механизм свертывания) на обнажившихся субэндотелиальных коллагеновых волокнах образуется комплекс из трех белков - фактора XII (фактор Хагемана), высокомолекулярного кининогена и прекалликреина. После связывания с высокомолекулярным кининогеном фактор XII медленно превращается в активную протеазу (фактор XIIа), которая, в свою очередь, превращает прекалликреин в калликреин, а фактор XI в активную форму (фактор ХIа).
Калликреин ускоряет превращение фактора XII в ХIIа, а фактор ХIа участвует в последующих реакциях свертывания.
Все эти взаимодействия хорошо изучены in vitro, однако могут существовать и иные механизмы активации фактора XI, так как у больных с дефицитом фактора XII, дефицитом высокомолекулярного кининогена и дефицитом прекалликреина гемостаз внешне нормален и кровоточивости нет.
В реакции 2 (внешний механизм свертывания крови) происходит превращение фактора VII в активную протеазу. При этом образуется комплекс, состоящий из фактора VII, Са2+ и тканевого фактора (липопротеида, который имеется во всех клеточных мембранах и обнажается после повреждения клетки).
Предполагается, что внешний механизм свертывания постоянно активен и обеспечивает фоновое свертывание крови.
В реакции 3 протеазы, образовавшиеся в предыдущих реакциях, активируют фактор X.
Первый способ состоит в образовании на фосфолипидных мембранах комплекса фактора VIII, фактора IX и фактора X с участием Са2+. Входящий в состав этого комплекса фактор IX активируется фактором ХIа, образовавшимся в реакции 1, и превращается в фактор IХа. Затем фактор IХа в комплексе с фактором VIII активирует фактор X.
Второй способ состоит в прямой активации факторов IX и X фактором VIIa, образовавшимся в реакции 2.
Активация факторов IX и X обеспечивает важную связь между внешним и внутренним механизмами гемостаза
В заключительной реакции 4 фактор Ха превращает протромбин в тромбин в присутствии фактора V, Са2+ и фосфолипидов. Хотя эта реакция может идти на любой (даже искусственной) фосфолипидной матрице, она протекает в несколько тысяч раз быстрее на поверхности активированных тромбоцитов.
Образующийся тромбин участвует сразу в нескольких процессах. Хотя его основная роль - превращение фибриногена в фибрин, он активирует также факторы V, VIII и XIII и, кроме того, стимулирует активацию и дегрануляцию тромбоцитов. После отщепления фибринопептида A и фибринопептида В от Aальфа- и Вбета-цепей фибриногена образуются фибрин-мономеры, которые полимеризуются в нити фибрина. Затем фактор ХIIIа (трансглутаминаза) образует между нитями фибрина поперечные сшивки, тем самым стабилизируя их (рис. 60.5).
Для действия фактора VII, фактора IX, фактора X и протромбина (фактор II) требуется Са2+. Кроме того, при синтезе всех этих белков в печени происходит присоединение второй карбоксильной группы (под действием витамин-К-зависимой гамма-глутамилкарбоксилазы) к некоторым остаткам глутаминовой кислоты с превращением ее в гамма-карбоксиглутаминовую кислоту. Пары карбоксильных групп связывают Са2+, в результате чего факторы свертывания закрепляются на отрицательно заряженных фосфолипидных мембранах и могут проявить свою активность.
Ингибирование этой посттрансляционной модификации лежит в основе действия непрямых антикоагулянтов (антагонистов витамина К).
Описанная классическая схема свертывания крови практически полезна, но оставляет без ответа ряд вопросов:
- почему дефицит фактора XII не сопровождается кровоточивостью, несмотря на резкое удлинение АЧТВ;
- почему у больных с дефицитом фактора XI кровоточивость выражена по-разному;
- почему дефицит фактора VIII и дефицит фактора IX (гемофилия A и гемофилия В) дает столь тяжелую кровоточивость, несмотря на сохранность внешнего механизма гемостаза?
Есть предположение, что основную роль в запуске свертывания играет активация фактора IX и фактора X комплексом тканевой фактор - фактор VIIa. После начала свертывания это взаимодействие блокируется ингибитором внешнего механизма свертывания, после чего дальнейшее образование тромбина зависит от внутреннего механизма и, в первую очередь, от фактора VIII и фактора IX.






