

История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
История развития хранилищ для нефти: Первые склады нефти появились в XVII веке. Они представляли собой землянные ямы-амбара глубиной 4…5 м...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Определение места расположения распределительного центра: Фирма реализует продукцию на рынках сбыта и имеет постоянных поставщиков в разных регионах. Увеличение объема продаж...
Интересное:
Берегоукрепление оползневых склонов: На прибрежных склонах основной причиной развития оползневых процессов является подмыв водами рек естественных склонов...
Уполаживание и террасирование склонов: Если глубина оврага более 5 м необходимо устройство берм. Варианты использования оврагов для градостроительных целей...
Влияние предпринимательской среды на эффективное функционирование предприятия: Предпринимательская среда – это совокупность внешних и внутренних факторов, оказывающих влияние на функционирование фирмы...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
Наиболее подробно у позвоночных изучено взаимодействие первичных афферентных волокон с мотонейронами спинного мозга. У круглоротых, как указывалось выше, часть клеток Рогон-Берда и клеток спинальных ганглиев не только устанавливает контакты с интернейронами, но и активирует мотонейроны посредством образования аксо-соматичеcких синапсов электро-химического типа.

Рис. 17. Окончание афферентных волокон в спинном мозге карпа (а) и на" одиночном мотонейроне спинного мозга лягушки (б) (Моторина, 1982; 1983).
1 - афферентные волокна и их ветви; 2 - мотонейроны; 3,4 - задние (3) и передние (4) рога серого вещества; 5, 6 - аксон (5) и дендрит (6) мотонейрона.
У рыб выявлены афферентные волокна двух типов. Одни из них оканчиваются на интернейронах медиальной части дорсального рога серого вещества спинного мозга, другие образуют аксо-соматические и аксо-дендритные контакты смешанного типа на мотонейронах вентрального рога. Часть афферентных волокон может переходить на противоположную сторону спинного мозга (рис. 17). У ряда изученных хрящевых рыб (например, некоторые скаты из группы Rajaformes и акулы Squalus) обнаружены прямые моносинаптические связи с мотонейронами, тогда как у других (например, акулы Scyliorhinus canicula) таких связей не выявлено.
У амфибий афферентные волокна образуют контакты не только с дистальными дендритами мотонейронов (как у большинства рыб), но и с их проксимальными ветвями и телами. Число контактов отдельного волокна с мотонейроном может достигать 60 - 70 (рис. 17). Показано, что афферентные волокна от определенной мышцы ветвятся среди большой группы мотонейронов, связанных с иннервацией различных мышц, но образуют контакты только с мотонейронами "своей" мышцы. У амфибий в спинном мозге, впервые среди позвоночных, намечается четкое топографическое разделение зон ветвления афферентов разного типа. Первичные афференты мышечных волокон (тип А) контактируют с мотонейронамн в вентральном. роге, тогда как афференты кожной и мышечной чувствительности (типы Iа, II) связаны с мотонейронами через систему интернейронов, и зона их ветвления ограничена дорсальным рогом.
|
|
Распределение афферентов в спинном мозге рептилий сходно с таковым у амфибий. Часть волокон оканчивается в дорсальном роге, другие достигают вентральных рогов. Электро-физиологически показано наличие прямых моносинаптических связей афферентов с мотонейронами у ящериц, крокодилов и черепах.
У млекопитающих и птиц происходит дальнейшая дифференциация входов. Основная масса проекций от кожных рецепторов (связанных с ноци-, механо- и терморецепцией) направляется в ядра дорсального рога серого вещества, а проекции таких специализированных рецепторов, как сухожильные, расположены в основании дорсального рога. Таким образом, большинство афферентов не достигает вентральных рогов спинного мозга, их контакт с мотонейронами осуществляется полисинаптически, через систему интернейронов (рис. 18). Ветвления отдельных афферентных волокон строго упорядочены и формируют сложную систему пространственных модулей, обеспечивающих высокий уровень специфичности межнейронных связей.
Серое вещество спинного мозга состоит из тел и отростков нейронов и у всех позвоночных расположено вокруг центрального спинномозгового канала, являющегося остатком полости
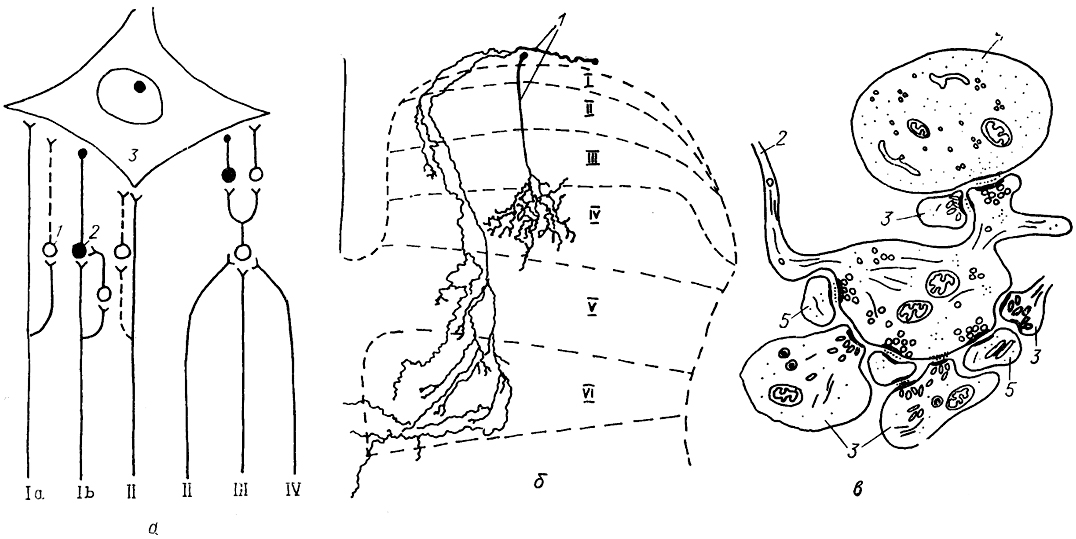
Рис. 18. Схема окончания первичных афферентов на мотонейроне (а), характер их ветвления в сером веществе задних рогов (б) и фрагмент ультраструктуры афферентного окончания (в) (Mannen, 1978; Honk, Zevrymer, 1981; Semba, 1983).
a: Ia, Ib, II-IV-различные виды афферентов; 1, 2 - возбуждающие (1) и тормозные (2) интернейроны; 3 - мотонейрон. б, в: 1 - афференты; 2, 3 - аксонные терминали афферентов типа Аα (2) и Аβ (3); 4, 5 - дендрит (4) и дендритный шипик (5) интернейронов задних рогов; I-VI - пластины Рекседа.
|
|
нервной трубки. Основная эволюционная тенденция в развитии серого вещества состоит, во-первых, в миграции клеток от центрального канала на периферию и, во-вторых, в прогрессивной дифференциации клеточных популяций на отдельные зоны и ядра. Так, у бесчерепных все клетки спинного мозга располагаются вокруг центрального канала, тогда как у круглоротых часть нейронов активно мигрирует в толщу спинного мозга, формируя две латерально расположенные группы. У остальных позвоночных в спинном мозге образуются отдельные популяции нейронов дорсальных и вентральных рогов. Позднее, у наземных позвоночных, в ряде отделов спинного мозга появляются боковые рога.
В 1952-1954 гг. Б. Рексед предложил новую универсальную классификацию серого вещества спинного мозга, разделив его на десять пластин (I-X), различающихся по структуре составляющих их элементов и по связям (рис. 15, а).
Пластина I представлена узкой полоской нейронов с характерными тангенциально ориентированными дендритами. На интернейронах пластины I оканчиваются афференты типов Аβ, Аδ и С. Аксоны интернейронов образуют восходящие тракты спинного мозга, а также посылают коллатерали в более глубокие слои спинного мозга (рис. 19). Помимо сенсорных волокон в I пластине обнаружены многочисленные афференты от вышележащих отделов спинного и головного мозга, в том числе от структур лимбической системы, через которые осуществляется контроль болевых ощущений.
Пластины II и III построены из нейронов среднего размера, дендриты которых могут проникать как в зону пластины I, так и в более глубокие слои серого вещества. Аксоны этих клеток распространяются на большие расстояния и захватывают несколько сегментов, образуя многочисленные внутри- и межсегментные связи. Часть аксонов образует систему восходящих трактов.
Обилие входящих в пластины I-III афферентных волокон, разнообразие нейронных типов, наличие синоптических комплексов конвергентного и дивергентного типов и модуляторных контактов определяют сложный характер межнейронных взаимодействий. Так, в вентральной части пластины III особенно многочисленны специфические формы аксо-аксональных синапсов между нейронами и афферентами, а также синапсы дендро-дендритного типа (рис. 18). Таким образом, значение нейронных комплексов пластин I-III состоит в контроле поступающей в спинной мозг сенсорной информации различного вида, в ее переработке и передаче в спинной и головной мозг.
|
|
Пластины IV-VI представлены скоплениями нейронов мультиполярного типа, дендриты которых ориентированы преимущественно в дорсальном направлении и проникают в зону пластин I-III. На телах и дендритах нейронов оканчиваются
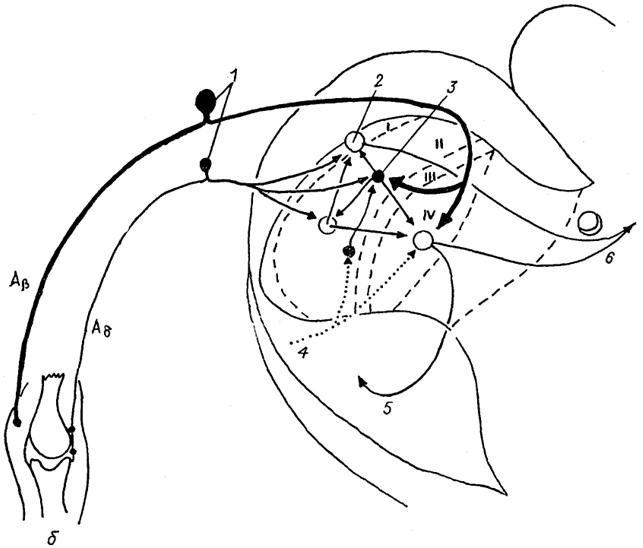
Рис. 19. Межнейронные связи в I-IV пластинах заднего рога спинного мозга (Delwaide, Schoenen, 1989).
Aβ, Aδ - афференты; 1 - чувствительные нейроны спинального ганглия; 2, 3 - возбуждающие (2) и тормозные (3) интернейроны; 4 - нисходящий бульбо-спинальный тракт; 5, 6 - восходящие спинно-ретикуло-таламический (5) и спинно-таламический (6) тракты.
многочисленные терминали первичных афферентов из групп Аα, Аβ, Аδ, аксоны интернейронов собственного и соседних сегментов, а также волокна нисходящих трактов. Окончания афферентов строго упорядочены: в вентральный отдел пластины V проецируются болевые и высокопороговые механорецепторы, в средний - рецепторы кожи, в дорсальный - низкопороговые: механорецепторы волосяных фолликулов. Эти отделы спинного мозга являются источником основной массы проприоспинальных (собственных) путей спинного мозга, образуя три системы связей: короткие - в пределах 1-2 сегментов, промежуточные- 4-12 сегментов, и Длинные. Последние представлены аксонами, идущими в составе тонкого пучка Голля в продолговатый мозг, в то время как их коллатерали достигают промежуточного мозга. Нейроны, аксоны которых образуют систему длинных связей, локализованы в основном в пластине IV шейных и пластинах V и VI пояснично-крестцовых сегментов спинного
мозга. Промежуточные и короткие проприоспинальные, пути связаны с проведением информации от рецепторов мышц, суставов, сухожилий. Часть волокон оканчивается на крупных нейронах ядра Кларка в пластине V, образующих в свою очередь крупный восходящий дорсальный спинно-мозжечковый тракт. Нейроны ядра Кларка получают проекции от афферентов флексорного рефлекса (FRA), связанных с иннервацией конечностей, причем разные афференты проецируются на различные клетки этого ядра.
|
|
Пластины VII и VIII, как полагают, не получают прямых афферентных проекций и занимают промежуточное положение между задними и передними рогами серого вещества спинного мозга. Здесь широко представлены терминальные ветвления проприо- и супраспинальных трактов. Аксоны интернейронов VII и VIII пластин оканчиваются на мотонейронах и образуют восходящие и нисходящие тракты различного типа. Одной из разновидностей этих интернейронов являются клетки Реншоу, получающие коллатерали аксонов мотонейронов и осуществляющие обратное торможение последних. Латеральные отделы пластины VII на уровне грудных - верхних поясничных сегментов образуют боковые рога и состоят из скоплений ассоциативных преганглионарных нейронов вегетативной нервной системы. Современные исследования, проведенные с помощью внутриклеточных инъекций пероксидазы хрена, показали сложную структурно-функциональную организацию боковых рогов. Их нейроны относятся к ретикулярному типу, для которого характерно широкое распространение дендритав, достигающих даже пластины I задних рогов. На дендритах и телах преганглионарных нейронов обнаружены многочисленные контакты различного происхождения. В целом подробный анализ нейронной структуры промежуточной зоны спинного мозга показал, что это образование ретикулярного типа, переходящее непосредственно в ретикулярную формацию ствола головного мозга и являющееся связующим звеном между сенсорными формациями заднего рога и моторными ядрами переднего рога.
Вентральные рога спинного мозга представляют особый интерес, так как здесь располагаются группы спинальных мотонейронов, управляющих деятельностью поперечнополосатой скелетной мускулатуры. В спинном мозге млекопитающих мотонейроны занимают район пластин VIII и IX и дифференцированы на группы ядер: медиальную, центральную и латеральную (рис. 15, а). Мотонейроны являются наиболее крупными клетками спинного мозга и разделяются на три группы: α-мотонейроны, крупные и обеспечивающие быстрые сокращения мышц; β-мотонейроны, мелкие и обеспечивающие медленные сокращения мышц, и γ-мотонейроны, связанные с иннервацией мышечных веретен. На примере организации моторных ядер наиболее отчетливо проявляется принцип соматотопии: каждая
мышца или группа мышц иннервируется определенной группой мотонейронов (мотонейронный пул). В пределах сегмента организация мотонейронных пулов млекопитающих подчиняется так называемому правилу Романеса: нейроны, иннервирующие дистальные мышечные группы конечностей, располагаются в спинном мозге дорсально по отношению к нейронам, иннервирующим проксимальные мышечные группы. При этом мотонейроны мышц-сгибателей и -разгибателей, часто относящихся к одной мышечной группе, в структуре моторных ядер также располагаются изолированно друг от друга.
Пластина X - центральное серое вещество спинного мозга занимает небольшой участок вокруг центрального канала. Здесь располагаются мелкие интернейроны, связывающие ее с другими участками серого вещества, а также проходят пучки комиссуральных волокон, соединяющих правую и левую половины спинного мозга.
|
|
|

Автоматическое растормаживание колес: Тормозные устройства колес предназначены для уменьшения длины пробега и улучшения маневрирования ВС при...

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

Историки об Елизавете Петровне: Елизавета попала между двумя встречными культурными течениями, воспитывалась среди новых европейских веяний и преданий...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!