Вторичный активный транспорт ионов
Помимо ионных насосов в клетках существуют системы, в которых перенос ионов через мембрану сопряжен не с гидролизом АТФ, а с работой окислительно-восстановительных ферментов. Транспорт веществ в этом случае является вторичным, опосредованным мембранным потенциалом и/ или градиентом концентрации ионов при наличии на мембране специфических переносчиков. Такой механизм переноса называют вторичным активным транспортом. В настоящее время известны три схемы вторичного активного транспорта: унипорт, антипорт, симпорт (рис. 10). На рисунке схематично изображен транспорт одновалентных ионов при помощи молекул-переносчиков. Подразумевается, что переносчик свободно пресекает мембрану, как в свободном, так и в связанном с ионом состоянии. Однонаправленный перенос иона в комплексе с переносчиком получил название унипорта. При этом через мембрану переносится заряд либо комплексом, если молекула переносчика электронейтральна, либо пустым переносчиком, если молекула переносчика заряжена. В результате такого транспорта происходит накопление ионов за счет снижения мембранного потенциала. Примером такого типа транспорта может служить накопление ионов калия в энергизированных митохондриях в присутствии валиномицина.

Рис. 10. Типы вторичного активного транспорта (Антонов, с.47).
Встречный перенос ионов одноместным переносчиком получил название антипорта. Перенос ионов осуществляется в два этапа: один ион в комплексе с переносчиком пересекает мембрану и комплекс распадается. Затем переносчик транспортирует другой ион в обратном направлении. Величина мембранного потенциала при этом остается постоянной. Движущей силой в этом случае является разность концентрации одного типа ионов. Классическим примером антипорта является перенос через плазмалемму ионов калия и водорода с участием молекулы нигерицина.
Совместный однонаправленный перенос ионов с участием двухместного переносчика называется симпортом. В этом случае, через мембрану курсируют две электронейтральные частицы: комплекс переносчика с катионом и анионом и свободный переносчик. Движущей силой при таком переносе может служить разность концентрации одного из ионов. Предполагают, что по такой схеме происходит транспорт аминокислот в клетку. Калий-натриевый насос создает начальный градиент концентрации ионов натрия, которые затем по схеме симпорта способствуют накоплению аминокислот в клетке.
Таким образом, в процессе жизнедеятельности границы клетки пересекают разнообразные вещества, потоки которых эффективно регулируются. Регуляция потоков осуществляет мембрана с встроенными в нее транспортными системами (ионные насосы, переносчики и высокоселективные ионные каналы). Особое значение имеют механизмы ионного сопряжения, получившие название вторичного транспорта. Как видно, сочетание активного транспорта веществ с явлениями диффузного переноса в клеточных мембранах обеспечивает жизнедеятельность клетки.
Потенциал покоя
Мембранный потенциал не возбужденных клеток называют потенциалом покоя. Такой термин дал впервые использовал Л. Гальвани, когда он обнаружил разность потенциалов на поверхности невозбужденной мышечной клетки. Значение потенциала покоя определяется разной концентрацией ионов по разные стороны мембраны и транспортом ионов через мембрану. Рассмотрим возникновение электрического потенциала на модельной схеме. Если сосуд разделим полупроницаемой мембраной и поместим в оба отсека раствор одного и того же и электролита (например, КС1) разной концентрации. Мембрана способна пропускать К+ и не пропускает Сl- (Рис. 7). Вследствие диффузии по градиенту концентрации через мембрану возникает поток заряженных частиц (ионов калия) и возникает электрический потенциал, который препятствует перемещению ионов через мембрану по грдиенту концентрации. Через определенное время устанавливается ионное равновесие на мембране, когда скорость перемещения ионов в разные стороны мембраны одинакова. При этом значения электрохимических потенциалов по разные стороны мембраны выравниваются: m вн = mнар.
Так как, m = m0 + RT lnС + ZFΔφ, то
RT lnС вн + ZFφвн = RT lnС нар + ZFφнар.
Активный транспорт веществ через мембраны.
Процессы активного транспорта играют большую роль в функционировании живых систем. За счет них в различных отсеках клетки создаются и поддерживаются градиенты физических и химических величин (концентраций веществ, электрических потенциалов). Благодаря процессам активного и пассивного транспорта обеспечивается поддержание стационарного состояния (гомеостаза) живой системы. Существование активного транспорта веществ через биологические мембраны впервые было показано в экспериментах Уссинга в 1949 году на примере переноса ионов натрия через кожу лягушки.
Как было отмечено, активный транспорт веществ не происходит самопроизвольно и требует затраты энергии. В биологических мембранах активный транспорт молекул и ионов сопряжен с гидролизом АТФ, т.е. энергия макроэргической химической связи используется для переноса вещества. Согласно современным представлениям, на плазматических и внутриклеточных мембранах функционируют специальные ионные насосы, перекачивающие ионы за счет свободной энергии гидролиза АТФ. В сущности эти насосы представляют собой комплексы интегральных белков-ферментов (транспортных АТФ-аз) с молекулами липидов мембран. К настоящему времени известны три типа электрогенных ионных насосов, осуществляющих активный перенос ионов через мембраны: К+, Na+- зависимая АТФ-аза плазмалеммы (К+, Na+- насос), Са2+- зависимая АТФ-аза (Са2+- насос), Н+-зависимая АТФ-аза мембран хлоропластов и митохондрий (Н+ -насос, или протонная помпа).
При работе К+, Na+- зависимой АТФ-азы, при гидролизе одной молекулы АТФ, в клетку переносится 2 иона калия и одновременно из клетки выкачиваются 3 иона натрия. При работе Са2+- насоса за счет энергии гидролиза АТФ транспортируются 2 иона кальция, а при работе протонной помпы – 2 иона водорода. Молекулярные механизмы работы ионных насосов пока еще до конца не разгаданы. Тем не менее, основные этапы работы этих сложных систем известны. Наиболее изученным в этом плане является К+, Na+- насос. Рассмотрим подробно структуру и работу этого насоса. Эта система отвечает за поддержание высокой концентрации ионов калия и низкой концентрации ионов натрия внутри клетки по сравнению с окружающей средой. Na+, К+ - зависимые АТФ-азы обнаружены в плазматических мембранах различных типов клеток, и их работа имеет огромное физиологическое значение. На работу этих насосов клетка в состоянии покоя затрачивает более 30 % синтезируемой АТФ. В частности, за счет работы этих насосов поддерживается электрическая возбудимость клеток, например нервных и мышечных клеток животных, создается движущая сила для транспорта через плазматическую мембрану сахаров, аминокислот и других соединений.
Функционирование К+, Na+- зависимой АТФ-азы впервые было показано английским исследователем Я. Скоу. Он обнаружил фермент, гидролизующий АТФ только при добавлении в среду К+, Na+ и Mg2+. Сейчас известно, что плазматическая Na+, К+ - зависимая АТФ-аза представляет собой тетрамерный белок α2 β2 с м.м. 270 кД. Большая α-субьединица с м.м. 95 кД представлена интегральным белком и содержит участок связывающий молекулу АТФ (Рис. 1.)..

Рис.1. Схематичное изображение Na+, К+ - зависимого насоса на плазматической мембране (стр. 187,костюк)
Меньшая β- субьединца имеет м.м. 40 кД и располагается на наружней части билипидного слоя. Она содержит углеводные группы Участок связывания АТФ обращена к цитоплазме, и только АТФ, находящаяся внутри клетки, служит эффективным субстратом для этой АТФ-азы. Сердечные глюкозиды (стероиды) ингибируют АТФ-азу только в том случае, если они находятся снаружи клетки.
При гидролизе одной молекулы АТФ происходит перенос трех ионов Na+ из клетки и двух ионов К+ в клетку.
Цикл работы насоса включает следующие основные этапы:
Связывание АТФ (фосфорилирование АТФ-азы) происходит только в присутствии Na+ и Mg2+:
E + ATФ Na+, Mg2+ ® Е -Р + АДФ
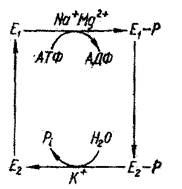
Рис. 2. Схема изменений конформации АТФ-азы в процессе транспорта ионов(стр.185, Костюк).
Далее в присутствии ионов калия происходит дефосфорилирование фермента:
Е -Р + Н2О К+® Е + Ф
Таким образом, мембранная АТФ-аза характеризуется Na+- - зависимым фосфорилированием и К+ –зависимым дефосфорилированием. Всего в процессе транспорта ионов калия и натрия и сопряженном с этим гидролизе молекулы АТФ фермент принимает, по крайней мере, четыре конформационные формы: Е1, Е1 – Р, Е2 – Р, Е2 (Рис. 9).
Е1 - свободная форма фермента, обладающая сродством к ионам натрия. Е1 – Р – первая фосфорилированная форма фермента в комплексе с 3 Na+. В этом состоянии ион связывающий участок обращен внутрь клетки и ионы Na+ захватываются из цитоплазмы клетки. Е2 – Р – вторая фосфорилированная форма фермента, в комплексе 2 ионами К+. В этом состоянии ион связывающий участок обращен к наружи клетки и ионы калия захватываются из внеклеточной среды. Е2 -свободная форма фермента, обладающая сродством к ионам калия. Таким образом, Na+ запускает фосфорилирование, а К+ - дефосфорилирование. Фосфорилирование стабилизирует форму Е2, дефосфорилирование – форму Е1 . Предполагается, что эти два формы фермента имеют небольшие конформационные различия.
Как видно из этой схемы, ключевыми этапами работы фермента являются: 1) образование комплекса фермента с АТФ на внутренней стороне мембраны (это реакция активируется ионами магния); 2) связывание фермент-субстратным комплексом трех ионов натрия; 3) фосфорилирование фермента с образованием АДФ; 4) конформационная перестройка (переворот «флип-флоп») фермента в мембране; 5) обмен ионов натрия на ионы калия, происходящая на внешней поверхности мембраны; 6) обратный переворот ферментного комплекса с переносом ионов калия внутрь клетки; 7) возвращение фермента в исходное состояние с освобождением ионов калия и неорганического фосфата.






