Введение
Физиология – это наука, предметом которой являются функции живого организма в условиях покоя и при различных видах деятельности, а также механизмы их регуляции.
Физиология, относясь к системе биологических наук, опирается на данные многих других биологических наук. Без знания морфологического строения клеток, тканей органов и систем органов, изучение их функций не может быть полноценным. Важнейшее значение имеет изучение физики и химии, так как все явления, происходящие в организме, обусловлены материальными процессами и в основе этих явлений лежат законы физико-химических наук. Развитие химии, физики и других смежных наук способствует более глубокому изучению многих физиологических закономерностей. Например, данные физики об электричестве послужили основой для создания нового раздела физиологии – электрофизиологии.
Большое значение для физиологии имеет развитие радиоэлектроники. Она дает возможность изучать физиологические процессы у свободно передвигающегося человека и животных, находящихся на расстоянии от исследователя.
Научно-технические достижения последних лет позволили обрабатывать экспериментальные физиологические материалы на электронно-вычислительных машинах, что позволило повысить точность выводов и дало возможность делать более широкие обобщения.
Функциональные изменения в живых организмах и механизмы их регуляции физиология изучает вместе с такими сопредельными науками, как биохимия и биофизика. Некоторые разделы физиологии, в особенности физиология нервной системы и анализаторов, тесно соприкасаются с психологией.
В ходе своего развития физиология прошла несколько этапов:
эмпирический, анатомо-функциональный, функциональный. На каждом этапе в изучении физиологического процесса или явления имело место два направления (подхода) - аналитическое и системное.
Аналитическое направление характеризуется изучением конкретного процесса, протекающего в каком-либо живом объекте (органе, ткани или клетке) как самостоятельного, т. е. вне связи его с другими процессами в изучаемом объекте. Такое направление дает всестороннее представление о механизмах данного процесса.
Системное направление ставит своей целью изучение конкретного процесса во взаимосвязи его с другими, протекающими на уровне организма как единого целого. Для физиологии как науки, необходимы оба эти направления. На разных этапах развития физиологии соотношение этих направлений изменялось: на ранних этапах развития физиологии преобладало аналитическое направление, на более поздних - системное. Для современного этапа характерно дальнейшее углубление аналитического подхода (изучение процессов на клеточном, субклеточном и молекулярном уровнях). Вместе с тем, стало обычным соотнесение этих процессов с процессами целого организма. Открытие системных закономерностей в деятельности живых организмов показало, что для выполнения определенных функций происходит избирательное объединение его отдельных органов и их систем, обеспечивающее достижение полезного приспособительного результата. Такие объединения были названы П. К. Анохиным функциональными системами. Применение системного подхода и, в частности, теории функциональных систем, дает возможность точнее и глубже понять закономерности спортивной деятельности.
Настоящее учебное пособие ставит главную задачу – способствовать упорядоченному усвоению материала, более эффективной самостоятельной работе студента по освоению курса физиологии. Дополнительным фактором, облегчающим самостоятельную познавательную деятельность, является приложение к пособию в виде мультимедийного диска с презентациями по рассматриваемым темам.
Рис. 1.1. Регистрация показателей физиологических систем.
Краткая история физиологии
Первые сведения о деятельности разных органов и систем организма были получены врачами Греции и Рима - Гиппократом, Аристотелем, Галеном. Эти сведения основывались на данных о строении тела, полученных при вскрытии трупов. Попытки изучения функций живого организма впервые были предприняты в начале нашей эры Галеном.
Началом современной физиологии как экспериментальной науки считают исследования, проведенные в начале XVII века английским врачом В.Гарвеем. Применив метод количественного изучения функций живого организма, он впервые описал движение крови по замкнутым сосудистым кругам. Был опубликован в 1628 году научный труд «Анатомические исследования движения сердца и крови у животных». Это первая работа по физиологии.
Большой вклад в понимание сущности реакций организма на раздражение внес в первой половине XVIII века физиолог и философ Рене Декарт. Он создал представления о путях, по которым проходит возбуждение в организме, обеспечивая ответные реакции на раздражения. Позднее на основе этих представлений чешский физиолог Иржи Прохазка (1749-1820) ввел сам термин «рефлекс», дал определение понятия «рефлекс» и в своем труде «Трактат о функциях нервной системы» заложил основы современной физиологии нервной системы.
Выдающимся достижением XVIII века явилось открытие биоэлектрических явлений, ("животного электричества") в 1791 г. итальянским анатомом и физиологом Луиджи Гальвани (Galvani, Luigi Aloisio, 1737-1798), что положило начало электрофизиологии.
Среди основоположников физиологии и экспериментальной медицины выдающееся место занимает немецкий естествоиспытатель Иоганнес Мюллер (Muller, Johannes Peter, 1801-1858), член Прусской (1834) и иностранный член-корреспондент Петербургской академии наук. Ему принадлежат фундаментальные исследования и открытия в области физиологии, патологической анатомии, эмбриологии. В 1833 г. он сформулировал основные положения рефлекторной теории, которые нашли дальнейшее развитие в трудах И.М.Сеченова и И.П.Павлова. И.Мюллер внес большой вклад в материалистическое познание природы. Он создал научную школу уникальную по количеству последователей и их вкладу в науку. К ней принадлежат Р.Вирхов, Г.Гельмгольц, Ф.Генле, Э.Дюбуа-Реймон, Э.Пфлюгер, Т.Шванн. В его лаборатории работали многие ученые России: А.М. Филомафитский, И.М.Сеченов и другие.
Значительного расцвета физиология достигла после великих открытий Шванна и Шлейдена (клеточная теория), Дарвина (эволюционное учение). В XIX и особенно XX веке физиология обогатилась новыми открытиями. Клод Бернар создал представление о гомеостазе, изучил роль нервной системы в регуляции тонуса сосудов и углеводного обмена. Дюбуа-Реймон явился основоположником электрофизиологии. Шеррингтон изучил физиологию спинного мозга. Капитальные исследования физиологии вегетативной нервной системы выполнил Кэннон.
 Физиологические исследования в России впервые были проведены в XVIII веке. Наибольшее значение среда них имеют исследования М.В.Ломоносова. Он сформулировал важнейший закон о сохранении вещества и энергии. Большой интерес представляют и его работы по физиологии органов чувств. В частности, он создал представлении о механизме цветного зрения.
Физиологические исследования в России впервые были проведены в XVIII веке. Наибольшее значение среда них имеют исследования М.В.Ломоносова. Он сформулировал важнейший закон о сохранении вещества и энергии. Большой интерес представляют и его работы по физиологии органов чувств. В частности, он создал представлении о механизме цветного зрения.
В России создание основ материалистического направления в физиологии прежде всего связано с деятельностью Алексея Матвеевича Филомафитского (1807-1849) - основоположника московской физиологической школы. В 1833 г. он защитил докторскую диссертацию "О дыхании птиц", затем в течение двух лет работал в Германии в лаборатории И.Мюллера. В 1835 г. А.М.Филомафитский стал профессором Московского университета, а в 1836 г. создал учебник "Физиология, изданная для руководства своих слушателей" (1836) - первый отечественный учебник физиологии. А.М.Филомафитский был одним из первых пропагандистов экспериментального метода в российской физиологии и медицине. Вместе с Н.И.Пироговым он разработал метод внутривенного наркоза, изучал вопросы физиологии дыхания, пищеварения, переливания крови ("Трактат о переливании крови", 1848); создал аппараты для переливания крови, маску для эфирного наркоза и другие физиологические приборы.
 Позднее, в 60-е годы XIX века, в России наблюдался значительный подъем физиологической мысли. Среди физиологов этого времени следует выделить И.М.Сеченова, которого И.П.Павлов назвал отцом русской физиологии. И.М.Сеченов впервые описал процессы торможения в центральной нервной системе. Особенное значение имеет разработанная им материалистическая теория психической деятельности человека. В своей работе «Рефлексы головного мозга» (1863 г.) он изложил свои взгляды на природу произвольных движений и психических явлений. И.М. Сеченов создал крупную физиологическую школу в России. Его учениками были Б.Ф.Вериго, Н.Е.Введенский, В.В.Пашутин, Г.В.Хлопин, М.Н.Шатерников и многие другие.
Позднее, в 60-е годы XIX века, в России наблюдался значительный подъем физиологической мысли. Среди физиологов этого времени следует выделить И.М.Сеченова, которого И.П.Павлов назвал отцом русской физиологии. И.М.Сеченов впервые описал процессы торможения в центральной нервной системе. Особенное значение имеет разработанная им материалистическая теория психической деятельности человека. В своей работе «Рефлексы головного мозга» (1863 г.) он изложил свои взгляды на природу произвольных движений и психических явлений. И.М. Сеченов создал крупную физиологическую школу в России. Его учениками были Б.Ф.Вериго, Н.Е.Введенский, В.В.Пашутин, Г.В.Хлопин, М.Н.Шатерников и многие другие.
 Огромное значение для развития физиологии имели работы И.П.Павлова и его учеников. И.П.Павлов используя методы хронических опытов, создал физиологию целостного организма. На первом этапе своей деятельности И.П.Павлов выполнил ряд важнейших исследований по физиологии кровообращения и пищеварения. В 1904 году И.П.Павлову за работы по физиологии пищеварения была присуждена Нобелевская премия. Следующий этап своих исследований И.П.Павлов посвятил изучению механизмов деятельности мозга, им было создано учение о высшей нервной деятельности, которое является естественнонаучной основой материалистического понимания природы сознания как высшей функции мозга человека. И.П.Павлов был учителем многих видных советских физиологов. Среди них следует назвать Л.А.Орбели - создателя эволюционной физиологии, разработчика многих вопросов, связанных с двигательной деятельностью. Учениками И.П.Павлова были также:
Огромное значение для развития физиологии имели работы И.П.Павлова и его учеников. И.П.Павлов используя методы хронических опытов, создал физиологию целостного организма. На первом этапе своей деятельности И.П.Павлов выполнил ряд важнейших исследований по физиологии кровообращения и пищеварения. В 1904 году И.П.Павлову за работы по физиологии пищеварения была присуждена Нобелевская премия. Следующий этап своих исследований И.П.Павлов посвятил изучению механизмов деятельности мозга, им было создано учение о высшей нервной деятельности, которое является естественнонаучной основой материалистического понимания природы сознания как высшей функции мозга человека. И.П.Павлов был учителем многих видных советских физиологов. Среди них следует назвать Л.А.Орбели - создателя эволюционной физиологии, разработчика многих вопросов, связанных с двигательной деятельностью. Учениками И.П.Павлова были также:
К.М.Быков - исследовал условнорефлекторную регуляцию деятельности внутренних органов;
Г.В.Фольборт - внесший вклад в изучение пищеварительных процессов и вопросов утомления и восстановления;
П.С.Купалов - изучавший различные закономерности условнорефлекторных реакций;
 П.К.Анохин (1898-1973) - выдвинул концепцию о системной деятельности нервной системы, предложил принцип опережающего отражения действительности, как базовый принцип живых организмов, создавший представления о функциональной системе, о системогенезе, давший начало многим направлениям современной физиологии, воспитавший ряд выдающихся физиологов.
П.К.Анохин (1898-1973) - выдвинул концепцию о системной деятельности нервной системы, предложил принцип опережающего отражения действительности, как базовый принцип живых организмов, создавший представления о функциональной системе, о системогенезе, давший начало многим направлениям современной физиологии, воспитавший ряд выдающихся физиологов.
Крупными представителями отечественной физиологии являются Н.Е.Введенский и А.А.Ухтомский. Н.Е. Введенский обнаружил в мышцах явления оптимума и пессимума, сформулировал понятие о лабильности нервов и мышц, создал учение о парабиозе. Дальнейшее развитие идеи Н.Е.Введенского получили в лабораториях его ученика А.А.Ухтомского, открывшего закон доминанты в деятельности нервной системы.
 Одним из крупнейших ученых, занимавшихся исследованием мозга, была Н.П.Бехтерева (1924 - 2008). Главными направлениями ее деятельности были работы по физиологии психической деятельности, структурно-функциональной организации и биоэлектрической активности головного мозга в норме и патологии.
Одним из крупнейших ученых, занимавшихся исследованием мозга, была Н.П.Бехтерева (1924 - 2008). Главными направлениями ее деятельности были работы по физиологии психической деятельности, структурно-функциональной организации и биоэлектрической активности головного мозга в норме и патологии.
П.В. Симонов (1926 - 2002) — советский, российский психофизиолог, биофизик и психолог. Главной сферой его научной деятельности была физиология высшей нервной деятельности, то есть изучение мозговых основ поведения. Им создан и экспериментально обоснован потребностно-информационный подход к анализу поведения и высших психических функций человека и животных, он внес крупнейший вклад в исследование эмоций человека.
Из советских физиологов также можно назвать ряд выдающихся ученых: Э.А.Асратян, А.Б.Коган, П.Г.Костюк, М.Е.Маршак, М.В.Сергиевский, В.Н.Черниговский, А.М.Уголев, и мн. др.
Вопросы для самоконтроля
Что изучает физиология?
Виды мышечных волокон
Мышцы состоят из мышечных волокон, обладающих разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов – гликолитических и окислительных. По соотношению миофибрилл, митохондрий и миоглобина различают так называемые белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна делят на быстрые, медленные и промежуточные. Если по активности АТФазы мышечные волокна различаются довольно резко, то степень активности дыхательных ферментов варьирует весьма значительно, поэтому наряду с белыми и красными существуют и промежуточные волокна.
Наиболее явно мышечные волокна различаются особенностями молекулярной организации миозина. Среди различных его изоформ существуют две основных – «быстрая» и «медленная». При постановке гистохимических реакций их различают по АТФазной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах (FF-волокна - быстро сокращающиеся, fast twitch fibres), преобладают гликолитические процессы, они более богаты гликогеном, в них меньше миоглобина, поэтому их называют также белыми. В медленных волокнах, обозначаемых как S (ST) волокна (slow twitch fibres), напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными. Они включаются при нагрузках в пределах 20-25% от максимальной силы и отличаются хорошей выносливостью.
 FT - волокна, обладающие по сравнению с красными волокнами небольшим содержанием миоглобина, характеризуются высокой сократительной скоростью и возможностью развивать большую силу. По сравнению с медленными волокнами они могут вдвое быстрее сокращаться и развить в 10 раз большую силу. FT-волокна, в свою очередь, подразделяются на FTO-и FTG-волокна. Существенные различия между перечисленными типами мышечных волокон определяется способом получения энергии (рис.2.1).
FT - волокна, обладающие по сравнению с красными волокнами небольшим содержанием миоглобина, характеризуются высокой сократительной скоростью и возможностью развивать большую силу. По сравнению с медленными волокнами они могут вдвое быстрее сокращаться и развить в 10 раз большую силу. FT-волокна, в свою очередь, подразделяются на FTO-и FTG-волокна. Существенные различия между перечисленными типами мышечных волокон определяется способом получения энергии (рис.2.1).
Рис. 2.1Различия энергообеспечения у мышечных волокон разных типов (по http://medi.ru/doc/g740203.htm).
Получение энергии в FTO-волокнах происходит так же, как и в ST-волокнах, преимущественно путем окислительного фосфорилирования. В связи с тем, что этот процесс разложения протекает относительно экономично (на каждую молекулу глюкозы при разложении мышечного гликогена для получения энергии накапливается 39 энергетических фосфатных соединений), FTO-волокна имеют также относительно высокую сопротивляемость утомляемости. Накопление энергии в FTG-волокнах происходит преимущественно путем гликолиза, т. е. глюкоза в отсутствии кислорода распадается до еще относительно богатого энергией лактата. В связи с тем, что этот процесс распада неэкономичен (на каждую молекулу глюкозы для получения энергии накапливается всего лишь 3 энергетических фосфатных соединения), FTG-волокна относительно быстро утомляются, но, тем не менее они способны развить большую силу и, как правило, включаются при субмаксимальных и максимальных мышечных сокращениях.
Двигательные единицы
Основным морфофункциональным элементом нервно-мышечного аппарата скелетных мышц является двигательная единица – ДЕ (рис.2.2.).

Рис 2.2. Двигательная единица
ДЕ включает мотонейрон спинного мозга с иннервируемыми его аксоном мышечными волокнами. Внутри мышцы этот аксон образует несколько концевых веточек. Каждая такая веточка образует контакт – нервно-мышечный синапс на отдельном мышечном волокне. Нервные импульсы, идущие от мотонейрона, вызывают сокращения определенной группы мышечных волокон. ДЕ мелких мышц, осуществляющих тонкие движения (мышцы глаза, кисти), содержат небольшое количество мышечных волокон. В крупных мышцах их в сотни раз больше.
ДЕ активизируются по закону „все или ничего". Таким образом, если от тела мотонейрона переднего рога спинного мозга посылается по нервным путям импульс, то на него реагируют или все мышечные волокна ДЕ, или ни одного. Для бицепса это означает следующее: при нервном импульсе необходимой силы укорачиваются все сократительные элементы (миофибриллы) всех (примерно 1500) мышечных волокон соответствующей ДЕ.
Все ДЕ в зависимости от функциональных особенностей делятся на 3 группы:
I. Медленные неутомляемые. Они образованы "красными" мышечными волокнами, в которых меньше миофибрилл. Скорость сокращения и сила этих волокон относительно небольшие, но они мало утомляемы, поэтому эти волокна относят к тоническим. Регуляция сокращений таких, волокон осуществляется небольшим количеством мотонейронов, аксоны которых имеют мало концевых веточек. Пример – камбаловидная мышца.
II В. Быстрые, легко утомляемые. Мышечные волокна содержат много миофибрилл и называются "белыми". Быстро сокращаются и развивают большую силу, но быстро утомляются. Поэтому их называют фазными. Мотонейроны этих ДЕ самые крупные, имеют толстый аксон с многочисленными концевыми веточками. Они генерируют нервные импульсы большой частоты. Напр., мышцы глаза.
II А. Быстрые, устойчивые к утомлению (промежуточные).
Все мышечные волокна одной ДЕ относятся к одному и тому же типу волокон (FT- или ST-волокна).
Мышцы, задействованные в выполнении очень точных и дифференцированных движений (например, мышцы глаз или пальцев руки), состоят обычно из большого количества ДЕ (от 1500 до 3000). Такие ДЕ имеют небольшое количество мышечных волокон (от 8 до 50). Мышцы, выполняющие относительно менее точные движения (например, большие мышцы конечностей), обладают существенно меньшим количеством ДЕ, но в их состав включено большое число волокон (от 600 до 2000).
В среднем человек имеет примерно 40% медленных и 60 % быстрых волокон. Но это средняя величина (по всей скелетной мускулатуре), мышцы же выполняют различные функции. Количественный и качественный состав мышц неоднороден, в них входит различное число двигательных единиц, соотношение типов которых также различно (композиция мышц). В связи с этим сократительные способности разных мышц неодинаковы. Наружные мышцы глаза, которые вращают глазное яблоко, развивают максимальное напряжение за одно сокращение длительностью всего 7.5 мс, камбалообразная – антигравитационная мышца нижней конечности, очень медленно развивает максимальное напряжение в течение 100 мс. Мышцы, выполняющие большую статическую работу (камбаловидная мышца), часто обладают большим количеством медленных ST-волокон, а мышцы, совершающие преимущественно динамические движения (бицепс), имеют большое количество FT-волокон.
Основные свойства мышечных волокон (следовательно, и двигательных единиц – ДЕ, в состав которых они входят), определяемые также свойствами мотонейронов, представлены в таблице 1.
Таблица 2.1
Композиция мышц
Соотношение типов мышечных волокон значительно различается у разных людей. Эти отличия видны у представителей разных спортивных специализаций (рис.2.3.).
Рис.2.3. Представленность быстрых (FF) и промежуточных (FR) волокон у спортсменов разных специализаций
У бегунов на длинные дистанции в икроножной мышце и пловцов-стайеров в дельтовидной мышце было обнаружено 90% медленных волокон, а у спринтеров в икроножной мышце до 90% быстрых волокон. Эти индивидуальные различия величины распределения волокон, скорее всего не объясняются тренировкой, они обусловлены генетически. Это подтверждается, в частности, тем, что, несмотря на гармоничное развитие скоростной силы рук и ног, боксер или фехтовальщик может, например, иметь чрезвычайно „быстрые ноги" и „медленные руки". Прирожденное количество быстрых FF-волокон является, видимо, причиной этого несоответствия. Тот факт, что у хороших представителей видов спорта, где особенно требуется выносливость (марафонцы, велосипедисты-шоссейники и т. д.), в основном преобладают медленные S-волокна, а высококлассные атлеты, которые демонстрируют скоростную силу (спринтеры, копьеметатели, толкатели ядра), обладают высоким процентом быстрых FF-волокон, свидетельствует об особом предрасположении именно к этим видам спорта.
То, что у элитных тяжелоатлетов соотношение FF-и S-волокон сбалансировано, связано со спецификой работы тяжелоатлета: опора и удержание в значительной степени выполняется посредством S-волокон.
Принадлежность мышечных волокон к определенной ДЕ задается от природы и не может быть изменена тренировкой. Каждый человек обладает индивидуальным набором S-и FF-волокон, количество которых, как показывают исследования, нельзя изменить при помощи специальной тренировки. Результаты исследований моно - (генетически идентичных) и ди-зиготных (генетически неидентичных) близнецов показывают, что у первых поразительно близко соотношение двух типов волокон в мышцах (даже если один из пары активно занимается спортом, а другой нет), тогда как у вторых возможны большие вариации в композиции мышцы.
Соответствующей силовой тренировкой можно относительно быстро преобразовывать FF-волокна в FR-волокна. Это дает возможность достигать хорошей выносливости даже тем спортсменам, которые, имея много быстрых FF-волокон, казалось бы, более подходят для проявления максимальной и скоростной силы. Несмотря на то, что тренировкой нельзя изменить унаследованное соотношение между S- и FF-волокнами, свойства волокон, хоть и в определенных пределах, все же приспосабливаются к предъявляемым специфическим раздражениям (поперечное сечение, время сокращения, оснащение энергоносителями и митохондриями и т. д.). В первую очередь это обусловлено тем, что целенаправленная тренировка на выносливость приводит к тому, что в FF волокнах количество митохондрий увеличивается в 3-4 раза.
У людей белые и красные волокна, как правило, перемешаны, но красные волокна преобладают в антигравитационных мышцах, а белые - в конечностях. Количество S и FF волокон в мышцах человека в среднем составляет 55 и 45 % соответственно.
Последовательность включения (рекрутирования) мышечных волокон в работу регулируется нервной системой и зависит от интенсивности нагрузки. При физической работе небольшой интенсивности (умеренной) – 20-25% уровня максимальной силы мышечного сокращения – в работу вовлекаются в основном медленные волокна. При работе нарастающей мощности последовательно включаются промежуточные, а затем и быстрые волокна. Однако даже при максимальной интенсивности в работу вовлекаются не все имеющиеся волокна: у нетренированных – не более 55-65 % имеющихся мышечных волокон, а у высоко тренированных спортсменов силовых видов спорта могут вовлекаться 80-90% ДЕ.
Природа возбуждения
Первые попытки последовательной разработки учения о "животном электричестве" связаны с именем Л. Гальвани. Э. Дюбуа-Реймон впервые показал, что наружная поверхность мышцы заряжена положительно по отношению к ее внутреннему содержимому. Следовательно, в состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая затем была названа мембранным потенциалом покоя или мембранным потенциалом. Его величина у разных клеток колеблется от 60 до 90 мВ.
А. Ходжкин, А. Хаксли и Б. Катц в 50-х годах 20 века объяснили причины возникновения мембранного потенциала покоя, для чего они существенно переработали ранее существовавшие представления и создали мембранно-ионную теорию. Согласно их взглядам мембранный потенциал покоя (МПП) обусловлен неодинаковой концентрацией ионов натрия, калия, кальция, хлора внутри клетки и во внеклеточной жидкости, а также неодинаковой проницаемостью для этих ионов поверхностной мембраны клетки (рис. 2.4). Цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Следовательно, в состоянии покоя существует асимметрия концентрации ионов внутри клетки и в окружающей ее среде.

Рис. 2.4. Регистрация потенциала покоя
В мембране имеются ионные каналы, образованные макромолекулами белка, пронизывающих липидный слой. Каналы мембраны делятся на неспецифические (каналы утечки) и специфические (селективные, обладающие способностью пропускать только определенные ионы). Неспецифические каналы пропускают различные ионы и открыты постоянно. Специфические каналы открываются и закрываются в ответ на изменения МПП. Эти каналы называются потенциалозависимыми.
В состоянии физиологического покоя мембрана нервных волокон в 25 раз лучше проницаема для K+, чем для Na+.
Выход положительно заряженных ионов калия приводит к появлению положительного заряда на наружной поверхности мембраны. Органические анионы - крупномолекулярные соединения, которые несут отрицательный заряд, и для которых мембрана клетки непроницаема, придают в этих условиях внутренней поверхности мембраны отрицательный заряд (рис.2.5).
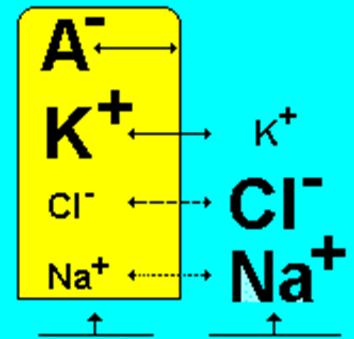
Рис.2.5. Концентрация основных ионов внутри и вне клетки.
В состоянии покоя существуют небольшие перемещения K+ и Na+ через мембрану по их концентрационному градиенту (табл.2.2), K+ больше, чем Na+.
Табл. 2.2.
Синхронизация работы ДЕ.
Так, если все 10 ДЕ начнут одновременно возбуждаться, то сила будет, например, 4 кгс/см2, а если они возбуждаются асинхронно, то максимальная сила составит 3 кгс/см2.
−Факторы межмышечной координации (мобилизация агонистов, торможение антагонистов, адаптационно-трофические влияния симпатической нервной системы - феномен Орбели - Гинецинского).
−Гормональные влияния (гормоны с анаболическим эффектом: половые, гормон роста и др.)
Для практических нужд важным является непосредственное измерение силовых показателей различных мышечных групп.
К Методам измерения мышечной силы относят:
1. Динамометрию
2. Динамографию
3. Электростимуляцию
4. Важными характеристиками силовых способностей являются время напряжения и расслабления, а также латентное время напряжения и расслабления.
Рис.2.11. Взаимное расположение волокон актина и миозина в различных участках саркомера.
Механизм мышечного сокращения
При световой микроскопии было замечено, что в момент сокращения ширина А-диска не уменьшается, а I-диски и Н-зоны саркомеров суживаются. Ряд фактов был установлен с помощью электронной микроскопии. Наиболее важно то, что длина нитей актина и миозина в момент сокращения не изменяется. Поэтому Хаксли и Хэнсон разработали теорию скольжения нитей.
Согласно этой теории мышца укорачивается в результате движения тонких актиновых нитей в промежутки между миозиновыми.
Это приводит к укорочению каждого саркомера, образующего миофибриллы. Скольжение же нитей обусловлено тем, что при переходе в активное состояние головки отростков миозина связываются с центрами актиновых нитей и вызывают их движение относительно себя ( рис.2.12).

Рис. 2.12. Скольжение актиновых и миозиновых нитей в процессе мышечного сокращения.
Сокращение начинается с того, что в области концевой пластинки двигательного нерва возникает потенциал действия (ПД). Он с большой скоростью распространяется по сарколемме и переходит с неё по системе поперечных трубочек саркоплазматического ретикулума (СР), на продольные трубочки и цистерны. Возникает деполяризация мембраны цистерн и из них в саркоплазму высвобождаются ионы кальция (Са2+). На нитях актина расположены молекулы еще двух белков - тропонина и тропомиозина. В состоянии покоя при низкой (менее 10-8 М) концентрации Са2+, тропомиозин блокирует присоединение мостиков миозина к нитям актина. Когда ионы Са2+ начинают выходить из СР (цистерн), молекула тропонина изменяет свою форму таким образом, что освобождаются головки миозина и начинается скольжение за счет ритмического замыкания и размыкания поперечных мостиков миозина с нитями актина. При этом головки ритмически продвигаются по нитям актина к Z- мембранам.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са2+-насос (фермент Са-АТФаза). Ионы Са2+ вновь закачиваются в цистерны саркоплазматического ретикулума, и их концентрация падает ниже 10-8 М. Молекулы тропонина приобретают исходную форму, и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от активных центров актина, и мышца за счет эластичности приходит в исходное расслабленное состояние.

Рис. 2.13. Механизм сопряжения возбуждения и сокращения (http://www.biobsu.org/phha/06/06_text.html).
Поперечная трубочка саркоплазматической мембраны, 2 –саркоплазматический ретикулум, 3 – ион Са2+, 4 – молекула тропонина, 5 – молекула тропомиозина
Существенную роль в сократительной активности мышц играют ионы Ca2+. Они активны в саркоплазме при очень низкой (10 - 6М и менее) концентрации (даже в воде двойной дистилляции концентрация Ca2+ выше, чем 10 - 6 М). Скелетная мышца не сокращается в ответ на деполяризацию мембраны, если исчерпаны запасы кальция во внутренних депо, а подвергнутые предварительной экстракции препараты волокон скелетной мышцы не сокращаются при добавлении АТФ, если отсутствует Са 2+).
Достаточно часто сравнивают процесс сократительного акта в саркомере с движениями гребцов в академической лодке. От филаментов миозина с обеих сторон, подобно веслам в лодке, отходят отростки - миозиновые мостики. В результате реакции на нервный сигнал эти мостики временно пристыковываются к филаментам актина (замыкание), а затем отводятся в позицию под углом 45". За счет этих движений филаменты актина перемещаются между филаментами миозина. После выполнения „гребка" отростки миозина, отрываются от актина и возвращаются в исходное положение (размыкание). За один цикл «замыкания - размыкания» саркомер укорачивается примерно на 1 % своей длины. Следовательно, для эффективного напряжения мышцы требуется большое число таких циклов соединения филаментов. В зависимости от типа мышечного волокна и требуемой величины напряжения мотонейроны генерируют импульсы с частотой от 7 до более чем 50 импульсов в секунду.
Саркомеры, расположенных по ходу миофибриллы, включается последовательно, в процессе работы мышцы их единичные минимальные сокращения суммируются, и миофибрилла сокращается на 25-30%. За счет плотной упаковки миофибрилл, их относительно небольшие сократительные силы складываются в суммарную силу мышечного волокна и в итоге в мышцы.
При непосредственном раздражении мышцы (прямое раздражение) или опосредовано через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение (рис.2.14), в котором выделяют три фазы:
• латентный период - время от начала действия раздражителя до начала ответной реакции;
• фаза сокращения (фаза укорочения);
• фаза расслабления.
В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например, так могут сокращаться быстрые глазодвигательные мышцы. Чаще всего одиночные сокращения суммируются.
Суммация - это сложение двух последовательных сокращений мышцы при нанесении на нее двух пороговых или сверхпороговых раздражений.
Различают 2 вида суммации: полную и неполную суммацию.
 Рис.2.14. Одиночное и тетаническое сокращение мышцы ( www.biobsu.org/phha/06/06_text.html ).
Рис.2.14. Одиночное и тетаническое сокращение мышцы ( www.biobsu.org/phha/06/06_text.html ).
Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу в начало фазы расслабления.
Полная суммация возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения.
Вопросы для самоконтроля
От чего зависит сила мышцы?
Центральная нервная система
Рис 3.1. Схема строения нейрона.
Каждая нервная клетка состоит из тела, множества коротких отростков – дендритов и чаще всего одного длинного отростка – аксона. Тело и дендриты играют основную роль в восприятии и переработке сигналов поступающих от других нейронов. Тело клетки выполняет питательную функцию. Функции аксона состоит в проведении нервных импульсов к другим нейронам или к исполнительным органам – мышцам, железам и т.п.
Передача возбуждения с одного нейрона на другой осуществляется при помощи синапса (рис. 3.2). Аксоны могут образовывать синапсы на теле другого нейрона или на его отростках.




Рис. 3.2. Строение синапса (http://ru.wikipedia.org/wiki/Синапс).
В наиболее общем (и весьма упрощенном виде) синапс имеет вид бляшки. Он состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны. В зависимости от механизма передачи нервного импульса различают химические и электрические синапсы.
В химическом синапсе внутри бляшек находятся пузырьки с медиатором (нейротрансмиттером ). Медиаторы – это биологически активные химические вещества, с помощью которых осуществляется передача электрического импульса с нервной клетки через синаптическое пространство между нейронами. По химической природе это аминокислоты, пептиды, моноамины (в том числе катехоламины). В настоящее время известно более 30 их разновидностей.
При достижении нервным импульсом пресинаптической мембраны происходит высвобождение медиатора в синаптическую щель. Диффундируя, медиатор попадает на постсинаптическую мембрану и, взаимодействуя с ней, изменяет проницаемость постсинаптической мембраны для ионов Na+ и K+, как это описано выше, в результате чего развивается деполяризации и возникает потенциал действия, распространяющийся по нейрону и его отросткам.
В эле







