Переваривание углеводов.
Это процесс гидролитического расщепления поли- и олигосахаридов. Переваривание начинается в ротовой полости. В слюне содержится фермент, расщепляющий крахмал (a-амилаза), расщепляет a-1,4-гликозидные связи внутри молекулы (но не концевые), при этом образуются декстрины, небольшое количество мальтозы и изомальтозы. В желудке углеводы не перевариваются, т.к. рН – кислая. Полное переваривание происходит в тонком кишечнике.
Поджелудочная железа вырабатывает a, b - амилазы и выделяет их в просвет кишечника. Клетки слизистой кишечника тоже продуцируют ряд ферментов, переваривающих олигосахара. Одна группа ферментов расщепляет a-гликозидные связи (мальтаза: a-1,4- гликозидные связи, изомальтаза-a-1,6; сахараза- сахарозу). Вторая группа расщепляет b-гликозидные связи (лактаза, b-галактозидаза, гетерогалактозидаза – расщепляет смешанные олигосахариды).
Под действием указанных ферментов происходит расщепление до моносахаров. Переваривание чаще идёт пристеночное (у поверхности клеток). Не перевариваются: клетчатка (нет фермента), но она нужна для перистальтики кишечника, пектозаны (полисахара, состоящие из пентоз). При переваривании больше всего образуется глюкозы. Всасывание осуществляется с помощью особых переносчиков, при этом характерна специфичность, затрачивается энергия (повышено потребление кислорода, расходуется АТФ).
Глюкоза и галактоза могут всасываться за счёт активного транспорта, за счёт грАДНента концентрации ионов Nа+, создаваемого Nа, К – АТФазой. Это обеспечивает их всасывание даже при низкой концентрации в кишечнике. С током крови по воротной вене глюкоза доставляется в печень, а частично с током крови доставляется во все органы и ткани, поступления и утилизации глюкозы в клетке. Глюкоза может поступить из ЖКТ с током крови, образоваться при распаде гликогена, может быть синтезирована из неуглеводных компонентов – глюконеогенез. Утилизация глюкозы:
Синтез гликогена, синтез жиров, синтез аминокислот, синтез других углеводов,полностью может расщепляться до СО2 и Н2О с образованием энергии.
Синтез и распад гликогена.
Гликоген является депо (запасом глюкозы). Гликоген образуется в печени и других тканях:

Гексокиназа малоспецифична, катализирует эту реакцию и с другими гексозами (фруктозой, маннозой и т.д.)
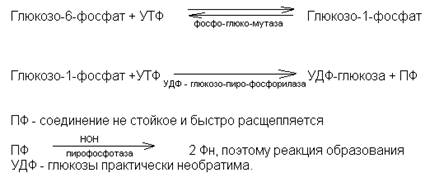

(порция гликогена) а)при образовании a-1,4-глюкогидных
n – не менее четырёх связей работает фермент; гликогенсинтаза
б)при образовании a-1,6-гликозидных
связей работает фермент; амило-1,4-1,6-
гликозилтрансфераза.
Ветвление цепи гликогена повышает его растворимость и, благодаря созданию большого количества не восстанавливающих концевых остатков, которые являются местами действия гликоген-синтазы, увеличивает скорость синтеза и расщепления гликогена.
По мере необходимости гликоген может расщепляться (и в печени, и в других тканях). Различают два пути расщепления гликогена:
Ø Амилолитический
Таким путём расщепляются боковые цепи – это около 10% гликогена. В тканях есть a-амилазы (расщепляют срединные a-1,4-гликозидные связи) и a-амилазы (расщепляют концевые a-1,4- гликозидные связи). По сути, идёт гидролиз гликогена:
Н2О
Гликоген → Глюкоза
Амилаза
Полученную глюкозу, в какие бы процессы она потом ни вступала (синтез, распад), необходимо проактивизировать, т.е. получить фосфориокислый эфир глюкозы, т.е. фосфорилировать её:
Глюкоза +АТФ → Глюкоза-6-Фосфат + АДФ
Как видно из приведённой реакции, мы затрачиваем при этом одну молекулу АТФ.
Ø Фосфоролитический (фосфоролиз):
Этим путём расщепляется до 90 % гликогена. Процесс выглядит по – другому:
1. Гликоген + Н3РО3 → Глюкозо-1-фосфат
фосфорилаза
2. Глюкозо-1-фосфат → Глюкозо-6-фосфат
фосфоглюкомутаза
Фосфорилаза расщепляет a-1,4- гликозидные связи, а a-1,6- гликозидные связи расщепляют a-1,6- гликозидазой. Но в реакции фосфоролиза участвует только активная форма – фосфорилаза «а», в то время, как в тканях вырабатывается и постоянно присутствует неактивная форма - фосфорилаза «b». Это впервые обнаружили в 1947 г. Дж. Корн, К. Корн. Превращение фосфорилазы «b» (неактивной) в фосфорилазу «а» (активную), происходит путём фосфорилирования. Фосфорилаза «b» представляет собой димер: каждая цепь содержит 841 АК, с молекулярной массой 100 000. При активации происходит фосфорилирование одного остатка серина (в 14 положении) в каждой субъединице.
 Фосфорилаза «b» + АТФ Фосфорилаза «а» + АДФ
Фосфорилаза «b» + АТФ Фосфорилаза «а» + АДФ
сер – ОН сер – О – РО3Н2
Этот процесс катализирует фермент: киназа фосфорилазы. (Активизирующее влияние на фосфорилазу «b» оказывает и АМФ (его много образуется при повышенном потреблении АТФ и, следовательно, возникает большая потребность в АТФ). АМФ оказывает активирующее влияние на фосфорилазу «b» вследствие аллостерического эффекта – путём связывания с регуляторным центром на каждой из субъединиц). Но этот фермент тоже вырабатывается неактивным, и также активизируется путём фосфорилирования. Однако частично активизировать киназу фосфорилазы могут и Са++ в концентрации 10-7м. Этот механизм активации имеет для организма важное значение, т.к. мышечное сокращение запускается высвобождением Са++-ионов. Таким образом, ращепление гликогена и мышечное сокращение связаны преходящим увеличением содержания Са++ в цитоплазме.
Фосфорилированию же киназы фосфорилазы способствует фермент протеинкиназа, которая активна лишь при присутствии в цитоплазме циклической формы АМФ.
АМФ образуется в плазматической мембране из АТФ:
 АТФ: Аденин-рибоза-Ф-Ф-Ф Аденин-рибоза- Ф + ПФ
АТФ: Аденин-рибоза-Ф-Ф-Ф Аденин-рибоза- Ф + ПФ
Аденилатциклаза (цАМФ)
Аденилатциклаза - мембраносвязанный фермент. На его активность в значительной степени влияют специфические гормоны.
Мы видим, что синтез и расщепление гликогена происходит разными путями. Главные ферменты синтеза и распада гликогена – это гликогенсинтеза и фосфорилаза. Синтез и расщепление гликогена координировано регулируются таким образом, что когда активируется гликогенсинтаза, то фосфорилаза остаётся почти неактивной и наоборот. На обмен гликогена большое влияние оказывают специфические гормоны:
инсулин (вырабатывается в поджелудочной железе (b-клетки)-повышай способность печени и мышц синтезировать гликoген (интимный механизм инсулина пока выяснен не в достаточной степени),Значительно лучше изучен механизм действия адреналина и глюкогона.
адреналин вырабатывается мозговым веществом надпочечников и вы-раженно стимулирует распад гликогена в мышцах и, в меньшей мере, в печени. Печень более чувствительна к гликогону, который вырабатывается a –клетками поджелудочной железы (при низком содержании глюкозы в крови).
Эрл Сазерленд установил, что действие адреналина и глюкогена на обмен углеводов опосредуется циклическим АМФ. Адреналин и глюкогон не проникают в свои клетки-мишени, они связываются с плазматическими мембранами и стимулируют аденилатциклазу.
Таким образом,получается следующая схема:
Адреналин (глюкагон)
Аденилатциклаза
 АТФ цАМФ
АТФ цАМФ
Протеинкиназа

Киназа фосфорилазы

 Фосфорилаза Фосфорилаза
Фосфорилаза Фосфорилаза
«b» «а»
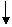
Фосфоролиз (распад гликогена)

Глюкозо-1-фосфат

Глюкозо-6-фосфат
Синтез гликогена тесно связан с его расщеплением. Активность гликоген-синтазы, подобно активности фосфорилазы, регулируется путем ковалентной модификации, т. е. фосфорилирования. В результате фосфорилирования гликоген-синтаза «а» (активная формула), превращается в неактивную форму-гликоген – синтазу «в».Фосфорилированная форма –гликоген-синтаза «в» требует для своей активности больших количеств глюкозо-6-фосфата.,тогда как дефосфорилированная «а» –форма активна и в присутствии и в отсутствии глюкозо-6-ффосфата. Таким образом,фосфорилирование оказывает противоположно направленное действие на активности гликоген-синтазы и гликоген-фосфорилазы. Так как фосфорилирование гликоген-синтазы «а» осущест-вляется протеинкиназой,то,когда протеин-киназа активна (см. схему, то-есть под действием адреналина и глюкогона), то она стимулирует выработку активной фосфорилазы «а» (через стАДНю киназы фосфорилазы) и неактивной гликоген-синтазы «в». Т. е. при повышенном распаде гликогена тормозится его синтез и наоборот.
Адреналин (глюкагон)
Аденилатциклаза
 АТФ цАМФ
АТФ цАМФ
 Протеникиназа Гликогенсинтаза «b»
Протеникиназа Гликогенсинтаза «b»
 фосфорилирование
фосфорилирование
 Cа++ Киназа фосфорилазы Гликогенсинтаза «а»
Cа++ Киназа фосфорилазы Гликогенсинтаза «а»

 Фосфорилаза «b» Фосфорилаза «а»
Фосфорилаза «b» Фосфорилаза «а»
| | | | | | |
| | |  |
| | |  |
| |
| | | Фосфоролиз гликогена гликогена
| |
| |
УДФ-глюкоза
 Глюкоза-1-фосфат Глюкоза-1-фосфат
Глюкоза-1-фосфат Глюкоза-1-фосфат
Глюкоза-6-фосфат Глюкоза-6-фосфат
СО2+ Н2О Глюкоза
Гликолиз.
Под гликолизом в общем смысле принято понимать разложение углеводов. В зависимости от конкретного организма и (или) условий его роста, гликолиз может идти по анаэробному пути (в этом случае он так и называется –анаэробный гликолиз).
И может идти по аэробному пути (соответственно – аэробный гликолиз). Для многих микроарганизмов, растущих в анаэробных условиях (в отсутствие кислорода) гликолиз является основным катаболическим путем для получения энергии из углеводных субстратов, приводя к образованию определенных конечных продуктов метаболизма, например, этанола, лактата, глицерина. Процесс такого типа часто называют брожением. У животных анаэробный гликолиз (с образованием лактата) обеспечивает энергией сокращение скелетных мышц в условиях ограниченного снабжения кислородом. В некоторых случиях гликолиз (аэробный гликолиз) идет по анаэробному пути в аэробных условиях т. е. конечным продуктом является лактат), хотя кислород и присутствует. Такие процес-сы идут, например, в эритроциттах, сетчатке, слизистой кишечника, в тканях плода (вскоре после рождения). Гликолиз –единственный анаэробный путь, дающий энергию. В аэробных условиях (в присутствии кислорода) реакции гликолиза составляют начальную форму разложения углеводов,связанную далее с циклом лимонной кислоты. В этом случае гликолиз останавливается на стАДНи образования ПВК. В аэробных условиях гликолиз и цикл Кребса совместно приводят к полному окислению гексозы до углекислого газа. Этот процесс сопровождается выделением больших количеств потенциально доступной метаболической энергии, в основном в виде высокоэнергетических продуктов НАДН и ФАДН2,которые, окисляясь в ходе дыхательной цепочки, дают энергию, необходимую для процесса ОФ, в результате которого образуется АТФ. Гликолиз –это первая до конца расшифрованная последовательность биохимических реакций, для этого потребовалось почти 50 лет (1890-1940). За это время были открыты фосфоэфиры, АТФ, НАД,выделены и охарактеризованы многие факторы и т.д.
Гликолиз протекает в цитозоле.
Схема гликолиза:
Очень важно образование на конечном этапе НАД+,который используется для окислительного фосфорилирования 3-фосфоглицеринового альдегида.
Лактат является конечным продуктом гликолиза. Т. к. лактат (как, впрочем, и пирува) достаточно легко проходит через плазматическую мембрану, он попадает из мышечной клетки (или эритроцитов) в кровь, с током крови попадает в печень, где окисляется в пируват, который в печени затем превращается в глюкозу. Таким образом образуется своего рода цикл. Эти превращения и составляют цикл Кори:
Вообще же в ходе гликолиза образуется 4 молекулы АТФ,а расходуется 2 АТФ. Таким образом, суммарный эффект окисления одной молекулы глюкозы в ходе гликолиза до лактата равен 2 АТФ. Утилизируется всего около 3% всей возможной энергии, которую можно было бы получать при окислении глюкозы, т.е. в сутки нужно было бы расщепить 27 кг глюкозы, чтобы обеспечить ортанизм энергией (если бы энергия получалась бы только путем гликолиза).
Однако,гликолиз имеет место в клетках. Он нужен в мышцах для быстрого получения энергии. А, например, эритророциты только за счет гликолиза и могут получать энергию, т. к. у них нет митохондрий, и, следовательно, нет и окислительного фосфорилирования. Большинство простых сахаров можно расщепить гликолитическим путем. Все, что для этого необходимо – это превратить эти сахара в Г-1- Ф, Г-6-Ф, Ф-6-Ф. Это происходит в ходе взаимного превращения моносахаров.
Аэробное окисление глюкозы
В аэробных условиях (в присутствии кислорода) реакции гликолиза составляет начальную фазу расщипления углеводов, связанную дальше с другим важным циклом- циклом лимонной кислоты (или его называют цикл трикарбоновых кислот, или цикл Кребса). В этом случае гликолиз останавливается на стАДНи образования пирувата, который затем путем окислительного декарбоксилирования превращается в ацетил- КоА, а ацетил-КоА, вступая в цикл Кребса, окисляется до СО2 и Н2О. Образующиеся в ходе цикла Кребса и в ходе окислительного декарбоксилирования восстановленные формы НАДН и ФАДН2 поступают в дыхательную цепь, где энергия, образующаяся при переносе с них Н+ и е- на О2 может быть использована, в ходе окислительного фосфорилирования, для синтеза АТФ. Легко рассчитать энергетический эффект этого процесса. На стАДНи образования пирувата из глюкозы образуются 2 молекулы АТФ и 2 молекулы НАДН2. На стАДНи образования ацетил-КоА из пирувата (в ходе окислительного декарбоксилирования пирувата) образуется 2НАДН2 (т.к. из глюкозы получается 2 молекулы пирувата). В ходе цикла Кребса образуется 1 моль ГТФ (=АТФ), 3 молекулы НАДН2 и 1 молекула ФАДН2 (все это умножает на 2, т. к. в цикл Кребса вступает 2 молекулы ацетил-КоА). Итого получается 2 молекулы АТФ + 2 молекулы ГТФ (=АТФ) + 10 молекул (2+2+6) НАДН и 2 моле-кулы ФАДН2. Одна НАДН при окислении в дыхательной цепи может дать энергию достаточную для синтеза 3-х АТФ, следовательно, 10 НАДН2 дадут 30 АТФ. Молекула ФАДН2, окисляясь в дыхательной цепи, может дать энергию, достаточную для синтеза 2 АТФ, а 2ФАДН2=4АТФ. Итого, получается 38 молекул АТФ. Всего при окислении одной молекулы глюкозы выделяется около 600 ккал. При аэробном окислении глюкозы, как мы подсчитали, образуется 38 молекул АТФ. АТФ, как известно, при гидролизе до АДФ дает = 10 ккал, т.е. всего на синтез АТФ при аэробном окислении затрачивается 380 ккал, что составляет около 60%,т.е. КПД аэробного окисления =60%, в то время, как КПД гликолиза составляет около 3 % (всего 2 молекулы АТФ).
Гликолиз протекает в цитозоле. Мы уже говорили о том, что ПВК относительно легко проходит через мембраны (когда говорим о судьбе лактата). Так и происходит, ведь все последующие этапы окисления глюкозы проходят в митохондриях. На начальных этапах окисления глюкозы (а они проходят в цитозоле) для окислительного фосфорилирования 3-фосфоглицеринового альдегида необходима окисленная форма НАД+. В ходе гликолиза она получается при восстановлении ПВК в лактат.При аэробном окислении глюкозы первый этап заканчивается образованием ПВК, которая через мембрану поступает внутрь митохондрий, подвергается окислительному декарбоксилированию, далее- цикл Кребса и дыхательная цепь. Можно было бы НАДН, образующуюся при фосфорилировании 3-фосфоглицеринового альдегида (т.к. мы не можем ее окислить с помощью ЛДГ- реакции до НАД+), отправить в митохондрии, там в дыхаательной цепи окислить до НАД+ и использовать для фосфорилирования 3-фосфоглицеринового альдегида. Но,оказалось, что НАДН и НАД+ не проходят через мембраны. Здесь действуют челночные механизмы: за счет НАДН образуется какая-то восстановленная форма вещества, которая проходит через мембраны, внутрь митохондрий, а окисленная форма возвращается в цитозоль, т.е. это вещество является, по сути, проводником протонов (переносчиком). При этом НАДН Окисляется до НАД+, что и требовалось. Так работает глицерофосфатный челночный механизм:
Так же работает и малатный (яблочная кислота (или малат)-ЩУК) челночный механизм. Мы уже говорили, что большие количества АТФ угнетают ключевые ферменты гликолиза. Т. к. аэробный распад глюкозы дает большой выход АТФ,. то включение этого процесса подавляет процесс распада углеводов в лактат (это эффект Пастера), т.е. гликолиз ингибируется при поступлении кислорода. Кроме того, что при накоплении АТФ ингибируются ферменты гликолиза (гексокиназа, фосфофруктокиназы и др.), есть и другие механизмы подавления гликолиза при поступлении кислорода. Например, в процессе гликолиза для образования АТФ путем фосфорилирования АДФ, который нужен и при дыхании (для окислительного фосфорилирования),т.е. возникает конкуренция, в которой выигрывают митохондрии. Для образования молочной кислоты из пирувата нужен НАДН, который также нужен и для работы челночных механизмов в аэробных условиях (для восстановления фосфодиоксиацетона до глицерофосфата), т.е.снова конкуренция и снова выигрывают митохондрии
Таким образом в аэробных условиях гликолиз не идет, хотя возможен (например, в опухолях, где угнетен фермент глицерофосфат-ДГ, который катализирует восстановление фосфодиоксиацетона до глицерофосфата- челноки не работают, идет гликолиз.
В скелетных мышцах, где снабжение кислородом невелико и метаболические процессы в большей степени протекают по анаэробному пути, АТФ поступает в результате гликолиза, так что основным субстратом служат запасы гликогена, а конечным продуктом – лактат. В других мышечных тканях, способных к аэробному метаболизму в жестких условиях (например, в сердечной мышце) в основном одновременно идет гликолиз с образованием пирувата и действует цикл лимонной кислоты. В этом случае основная часть необходимого АТФ доставляется благодаря процессу ОФ в митохондриях. В соответствии с такими функциональными различиями между аэробным и анаэробными мышечными тканями сердечная мышца обычно содержит гораздо больше митохондрий, чем клетки скелетных мышц. Около 40% Сухой массы сердца приходится на митохондрии.
 В периоды длительной мышечной активности АТФ используется непосредственно после его образования. В периоды отдыха энергия АТФ сохраняется в виде макророэрга – креатинфосфата, который в короткие периоды активности и в начальной фазе длительной активности служит донором фосфата для синтеза АТФ из АДФ.
В периоды длительной мышечной активности АТФ используется непосредственно после его образования. В периоды отдыха энергия АТФ сохраняется в виде макророэрга – креатинфосфата, который в короткие периоды активности и в начальной фазе длительной активности служит донором фосфата для синтеза АТФ из АДФ.
Далее фосфоенолпируват выходит в цитозоль и там продолжаются процессы, обратные гликолизу, т.е. продолжается глюконеогенез. Биотин всегда участвует в реакциях карбоксилирования. Обнаружили его случайно: крысам скармливали много куриного белка – крысы заболевали (дерматит, и т.д.). Оказалось, что в состав куриного белка входит белок авидин, который образует прочный комплекс с витамином Н, биотин не всасывается, в организме образуется его дефицит и, как следствие, заболевание.
Иногда карбоксикиназа (фермент, который катализирует переход ЩУК в фосфоенолпируват) обнаруживается в цитоплазме. Но метохондриальная мембрана непроницаема для ЩУК. Поэтому для третьего пути нужны ещё два фермента – малатдегидрогеназа (в митохондриях) и малат-ДГ (в цитоплазме). По этой схеме ЩУК в митохондриях восстанавливается в малат:
А вот яблочная кислота легко проходит через митоидриальную мембрану, где цитоплазматическая малат-ДГ окисляет её до ЩУК, а на ЩУК уже действует найденная ранее в цитоплазме фосфоенолпируваткарбоксикиназа, при этом ЩУК превращается в фосфоенол ПВК. И т.д. до образования глюкозы.
Биосинтез лактозы.
Синтез лактозы катализируется ферментом лактозо-синтазой. Это фермент состоит из 2-х субъединиц: каталитической (ее называют галактозилттрансфера-зой) и модифицирующей (ее называют галактозил трансферазой a - лактальбумин). Галактозилтрансфераза присутствует в большинстве тканей, где она участвует в синтезе углеводного компанента гликопротеинов. А вот лактозо-синтаза обнаружена только в молочной железе. Во время беременности галактозил-трансфераза интезируется и накапливается в молочной железе. А a-лактальбумин-синтезируется в неболь-шом количестве. При родах резкое изменение в содержании некоторых гормонов вызывают синтез больших количеств a-лактальбумина. Образующийся лактозо-синтазный комплекс синтезирует большие количества лактозы.
Переваривание углеводов.
Это процесс гидролитического расщепления поли- и олигосахаридов. Переваривание начинается в ротовой полости. В слюне содержится фермент, расщепляющий крахмал (a-амилаза), расщепляет a-1,4-гликозидные связи внутри молекулы (но не концевые), при этом образуются декстрины, небольшое количество мальтозы и изомальтозы. В желудке углеводы не перевариваются, т.к. рН – кислая. Полное переваривание происходит в тонком кишечнике.
Поджелудочная железа вырабатывает a, b - амилазы и выделяет их в просвет кишечника. Клетки слизистой кишечника тоже продуцируют ряд ферментов, переваривающих олигосахара. Одна группа ферментов расщепляет a-гликозидные связи (мальтаза: a-1,4- гликозидные связи, изомальтаза-a-1,6; сахараза- сахарозу). Вторая группа расщепляет b-гликозидные связи (лактаза, b-галактозидаза, гетерогалактозидаза – расщепляет смешанные олигосахариды).
Под действием указанных ферментов происходит расщепление до моносахаров. Переваривание чаще идёт пристеночное (у поверхности клеток). Не перевариваются: клетчатка (нет фермента), но она нужна для перистальтики кишечника, пектозаны (полисахара, состоящие из пентоз). При переваривании больше всего образуется глюкозы. Всасывание осуществляется с помощью особых переносчиков, при этом характерна специфичность, затрачивается энергия (повышено потребление кислорода, расходуется АТФ).
Глюкоза и галактоза могут всасываться за счёт активного транспорта, за счёт грАДНента концентрации ионов Nа+, создаваемого Nа, К – АТФазой. Это обеспечивает их всасывание даже при низкой концентрации в кишечнике. С током крови по воротной вене глюкоза доставляется в печень, а частично с током крови доставляется во все органы и ткани, поступления и утилизации глюкозы в клетке. Глюкоза может поступить из ЖКТ с током крови, образоваться при распаде гликогена, может быть синтезирована из неуглеводных компонентов – глюконеогенез. Утилизация глюкозы:
Синтез гликогена, синтез жиров, синтез аминокислот, синтез других углеводов,полностью может расщепляться до СО2 и Н2О с образованием энергии.
Синтез и распад гликогена.
Гликоген является депо (запасом глюкозы). Гликоген образуется в печени и других тканях:
Гексокиназа малоспецифична, катализирует эту реакцию и с другими гексозами (фруктозой, маннозой и т.д.)
(порция гликогена) а)при образовании a-1,4-глюкогидных
n – не менее четырёх связей работает фермент; гликогенсинтаза
б)при образовании a-1,6-гликозидных
связей работает фермент; амило-1,4-1,6-
гликозилтрансфераза.
Ветвление цепи гликогена повышает его растворимость и, благодаря созданию большого количества не восстанавливающих концевых остатков, которые являются местами действия гликоген-синтазы, увеличивает скорость синтеза и расщепления гликогена.
По мере необходимости гликоген может расщепляться (и в печени, и в других тканях). Различают два пути расщепления гликогена:
Ø Амилолитический
Таким путём расщепляются боковые цепи – это около 10% гликогена. В тканях есть a-амилазы (расщепляют срединные a-1,4-гликозидные связи) и a-амилазы (расщепляют концевые a-1,4- гликозидные связи). По сути, идёт гидролиз гликогена:
Н2О
Гликоген → Глюкоза
Амилаза
Полученную глюкозу, в какие бы процессы она потом ни вступала (синтез, распад), необходимо проактивизировать, т.е. получить фосфориокислый эфир глюкозы, т.е. фосфорилировать её:
Глюкоза +АТФ → Глюкоза-6-Фосфат + АДФ
Как видно из приведённой реакции, мы затрачиваем при этом одну молекулу АТФ.
Ø Фосфоролитический (фосфоролиз):
Этим путём расщепляется до 90 % гликогена. Процесс выглядит по – другому:
1. Гликоген + Н3РО3 → Глюкозо-1-фосфат
фосфорилаза
2. Глюкозо-1-фосфат → Глюкозо-6-фосфат
фосфоглюкомутаза
Фосфорилаза расщепляет a-1,4- гликозидные связи, а a-1,6- гликозидные связи расщепляют a-1,6- гликозидазой. Но в реакции фосфоролиза участвует только активная форма – фосфорилаза «а», в то время, как в тканях вырабатывается и постоянно присутствует неактивная форма - фосфорилаза «b». Это впервые обнаружили в 1947 г. Дж. Корн, К. Корн. Превращение фосфорилазы «b» (неактивной) в фосфорилазу «а» (активную), происходит путём фосфорилирования. Фосфорилаза «b» представляет собой димер: каждая цепь содержит 841 АК, с молекулярной массой 100 000. При активации происходит фосфорилирование одного остатка серина (в 14 положении) в каждой субъединице.
Фосфорилаза «b» + АТФ Фосфорилаза «а» + АДФ
сер – ОН сер – О – РО3Н2
Этот процесс катализирует фермент: киназа фосфорилазы. (Активизирующее влияние на фосфорилазу «b» оказывает и АМФ (его много образуется при повышенном потреблении АТФ и, следовательно, возникает большая потребность в АТФ). АМФ оказывает активирующее влияние на фосфорилазу «b» вследствие аллостерического эффекта – путём связывания с регуляторным центром на каждой из субъединиц). Но этот фермент тоже вырабатывается неактивным, и также активизируется путём фосфорилирования. Однако частично активизировать киназу фосфорилазы могут и Са++ в концентрации 10-7м. Этот механизм активации имеет для организма важное значение, т.к. мышечное сокращение запускается высвобождением Са++-ионов. Таким образом, ращепление гликогена и мышечное сокращение связаны преходящим увеличением содержания Са++ в цитоплазме.
Фосфорилированию же киназы фосфорилазы способствует фермент протеинкиназа, которая активна лишь при присутствии в цитоплазме циклической формы АМФ.
АМФ образуется в плазматической мембране из АТФ:
АТФ: Аденин-рибоза-Ф-Ф-Ф Аденин-рибоза- Ф + ПФ
Аденилатциклаза (цАМФ)
Аденилатциклаза - мембраносвязанный фермент. На его активность в значительной степени влияют специфические гормоны.
Мы видим, что синтез и расщепление гликогена происходит разными путями. Главные ферменты синтеза и распада гликогена – это гликогенсинтеза и фосфорилаза. Синтез и расщепление гликогена координировано регулируются таким образом, что когда активируется гликогенсинтаза, то фосфорилаза остаётся почти неактивной и наоборот. На обмен гликогена большое влияние оказывают специфические гормоны:
инсулин (вырабатывается в поджелудочной железе (b-клетки)-повышай способность печени и мышц синтезировать гликoген (интимный механизм инсулина пока выяснен не в достаточной степени),Значительно лучше изучен механизм действия адреналина и глюкогона.
адреналин вырабатывается мозговым веществом надпочечников и вы-раженно стимулирует распад гликогена в мышцах и, в меньшей мере, в печени. Печень более чувствительна к гликогону, который вырабатывается a –клетками поджелудочной железы (при низком содержании глюкозы в крови).
Эрл Сазерленд установил, что действие адреналина и глюкогена на обмен углеводов опосредуется циклическим АМФ. Адреналин и глюкогон не проникают в свои клетки-мишени, они связываются с плазматическими мембранами и стимулируют аденилатциклазу.
Таким образом,получается следующая схема:
Адреналин (глюкагон)
Аденилатциклаза
АТФ цАМФ
Протеинкиназа
Киназа фосфорилазы
Фосфорилаза Фосфорилаза
«b» «а»
Фосфоролиз (распад гликогена)
Глюкозо-1-фосфат
Глюкозо-6-фосфат
Синтез гликогена тесно связан с его расщеплением. Активность гликоген-синтазы, подобно активности фосфорилазы, регулируется путем ковалентной модификации, т. е. фосфорилирования. В результате фосфорилирования гликоген-синтаза «а» (активная формула), превращается в неактивную форму-гликоген – синтазу «в».Фосфорилированная форма –гликоген-синтаза «в» требует для своей активности больших количеств глюкозо-6-фосфата.,тогда как дефосфорилированная «а» –форма активна и в присутствии и в отсутствии глюкозо-6-ффосфата. Таким образом,фосфорилирование оказывает противоположно направленное действие на активности гликоген-синтазы и гликоген-фосфорилазы. Так как фосфорилирование гликоген-синтазы «а» осущест-вляется протеинкиназой,то,когда протеин-киназа активна (см. схему, то-есть под действием адреналина и глюкогона), то она стимулирует выработку активной фосфорилазы «а» (через стАДНю киназы фосфорилазы) и неактивной гликоген-синтазы «в». Т. е. при повышенном распаде гликогена тормозится его синтез и наоборот.
Адреналин (глюкагон)
Аденилатциклаза
АТФ цАМФ
Протеникиназа Гликогенсинтаза «b»
фосфорилирование
Cа++ Киназа фосфорилазы Гликогенсинтаза «а»
Фосфорилаза «b» Фосфорилаза «а»
| | | | | | |
| | | |
| | | |
| |
| | | Фосфоролиз гликогена гликогена
| |
| |
УДФ-глюкоза
Глюкоза-1-фосфат Глюкоза-1-фосфат
Глюкоза-6-фосфат Глюкоза-6-фосфат
СО2+ Н2О Глюкоза
Гликолиз.
Под гликолизом в общем смысле принято понимать разложение углеводов. В зависимости от конкретного организма и (или) условий его роста, гликолиз может идти по анаэробному пути (в этом случае он так и называется –анаэробный гликолиз).
И может идти по аэробному пути (соответственно – аэробный гликолиз). Для многих микроарганизмов, растущих в анаэробных условиях (в отсутствие кислорода) гликолиз является основным катаболическим путем для получения энергии из углеводных субстратов, приводя к образованию определенных конечных продуктов метаб