Тема «ГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ФУНКЦИЙ.
Транспорт либеринов и статинов.
Из нейронов они секретируются в кровь фенестрированных капилляров срединного возвышения (воронки) гипоталамуса. Переносятся с кровью к аденогипофизу и поступают через сеть капилляров к эндокринным клеткам аденогипофиза. (Двойная сеть капилляров препятствует поступлению этих гормонов в системный кровоток.) В аденогипофизе либерины и статины регулируют образование тройных гормонов (АКТГ, ТТГ, ФСГ, ЛГ, СТГ, пролактина, МСГ), изменяя порог их реакции на гормоны периферических желез.
Тропные гормоны аденогипофиза действуют на зависимые железы и ткани.
Саморегуляция активности гипоталамо-гипофизарной системы осуществляется с помощью отрицательных обратных связей.
Главное значение имеет длинная связь: влияние гормона периферической железы на продукцию тропных гормонов аденогипофиза. При этом порог реакции клеток аденогипофиза регулируется либеринами и статинами
Допускается наличие коротких связей: влияние тропных гормонов аденогипофиза на продукцию либеринов и статинов гипоталамуса.
Трансгипофизарная регуляция через нейрогипофиз.
В супраоптических и паравентрикулярных ядрах гипоталамуса образуются АДГ (вазопрессин) и ОКСИТОЦИН. (В гликопептиде-предшественнике они находятся вместе.)
В результате аксонного транспорта эти гормоны поступают в нейрогипофиз и депонируются в нем (окончания аксонов расположены на базальной мембране капилляров).
Из нейрогипофиза АДГ и окситоцин секретируются в околокапиллярное пространство и затем поступают в кровь, а также через Ш желудочек мозга в цереброспинальную жидкость (влияние на поведение и память).
Парагипофизарный путь.
Прямой нервный контроль секреции гормонов осуществляется в мозговом слое надпочечников, эпифизе, гипоталамусе (его нейросекреторных ядрах); в этих железах происходит преобразование нервного импульса в нейросекреторный процесс.
Эндокринная (негипофизарная) регуляция осуществляется в результате влияния гормонов периферических эндокринных желез друг на друга (например, торможение кортизолом секреции тироксина, соматостатином секреции инсулина и глюкагона).
Гуморальная регуляция секреции инсулина, альдостерона, паратгормона осуществляется соответственно уровнем глюкозы, Na+ и Са2+ в крови.
Методы исследования желез внутренней секреции.
- Удаление железы или подавление ее активности.
- Пересадка железы или введение гормонов.
- Определение количества гормонов и их метаболитов в биологических жидкостях (крови, моче, ликворе) биохимическими, иммунорадиологическими и имму-нофлюоресцентными методами анализа.
- Функциональные нагрузочные пробы (например, тест на толерантность к глюкозе).
Физиология гипофиза.
Гормоны нейрогипофиза.
Антидиуретический гормон (АДГ) или вазопрессин (нонапептид супраоптических и паравентрикулярных ядер гипоталамуса, концентрация в плазме 1-13 нг/л).
Эффекты АДГ (вазопрессина).
Увеличивает реабсорбцию воды в дистальных канальцах и собирательных трубочках почек, что приводит к уменьшению диуреза (эффект реализуется через У2-рецепторы, повышение уровня цАМФ и увеличение числа водных каналов в клеточной мембране).
В более высоких концентрациях усиливает сокращения гладких мышц и вызывает сужение сосудов.
Активирует центр жажды и питьевое поведение (одновременно тормозит секрецию ренина).
Участвует в механизмах формирования памяти (стимулирует обучение).
Регуляция секреции АДГ.
Главным регулятором секреции АДГ является осмоляльность плазмы, которая контролируется осморецепторами гипоталамуса, сосудов, тканей. Гиперосмия (>290 мосм/кг) стимулирует секрецию АДГ, гипоосмия (< 280 мосм/кг) тормозит секрецию АДГ.
Второй регулятор секреции АДГ - колебания объема циркулирующей крови (ОЦК) и АД, которые регистрируются барорецепторами аорты, сонной артерии, легочной артерии, волюморецепторами предсердий и вен. Гиповолемия (снижение ОЦК более чем на 5 %) и снижение АД стимулируют секрецию, гиперволемия (увеличение ОЦК) и повышение АД тормозят секрецию АДГ.
Другие факторы, влияющие на секрецию АДГ.
Стимулируют секрецию АДГ симпатоадреналовая система (через Р-адрено-рецепторы), ангиотензин II, гистамин, ацетилхолин.
Тормозят секрецию АДГ натрийуретический гормон предсердий, этанол, переохлаждение.
Окситоцин (образуется в паравентрикулярных и супраоптических ядрах гипоталамуса, концентрация в плазме 3,2 мМЕ/л).
Эффекты окситоцина реализуются через поступление Са +:
Усиливает сократительную активность матки во время родов (эстрогены увеличивают чувствительность матки к окситоцину, а прогестерон снижает).
Усиливает сокращение гладкой мускулатуры протоков молочных желез и выделение молока.
Стимулирует продукцию простагландинов в эндометрии, которые совместно с окситоцином вызывают начало родов (мобилизуют Са+).
Участвует в механизмах формирования памяти (нарушает запоминание).
Регуляция секреции окситоцина: рефлекторная стимуляция секреции с
рецепторов растяжения шейки матки и влагалища во время родов, сосков молочных желез при кормлении грудью; условнорефлекторная – на крик ребенка и кормление.
Околощитовидные железы.
Физиологические эффекты паратгормона - повышает уровень Са2+ в крови в результате действия следующих механизмов.
· Повышение активности остеоцитов и остеокластов (усиление резорбции костной ткани).
· Увеличение реабсорбции Са2+ в дистальных почечных канальцах и снижения реабсорбции фосфатов в проксимальных почечных канальцах (фосфатурия).
· Усиление всасывания Са2+ в тонком кишечнике (действует совместно с витамином D3).
Эффект паратгормона связан со стимуляцией аденилатциклазы, увеличением уровня цАМФ и активности протеинфосфокиназы А в клетках-мишенях.
Регуляция продукции паратгормона (суточный ритм секреции: в ночные часы в 2 - 3раза больше, чем в дневное время). Основной стимулятор секреции - гипокальциемия (ускоряет трансформацию пропаратгормона в активный гормон). Ингибитор секреции - снижение уровня Mg2+ в крови.
Эндокринные функции островков Лангерганса поджелудочной железы: α-клетки
(25 %) продуцируют глюкагон; Р-клетки (60 %) - инсулин; β-клетки (10 %) - соматостатин (все поли пептиды).
Эндокринные функции β-клеток, вырабатывающих инсулин (в крови находится в
свободной форме, концентрация в плазме 6-24 мМе/л).
Секреция инсулина стимулируется глюкозой крови, которая проникает в β-клетки преимущественно с помощью транспортера ГЛЮТ 2, что приводит к повышению уровня АТФ и закрытию АТФ-зависимьгх К+ - каналов. Возникает деполяризация мембраны до КУД и возникникает ПД. Вход Са2+ в клетку во время ПД запускает экзоцитоз гранул инсулина и открытие К+-каналов. Секреция инсулина осуществляется в кровь панкреатической вены.
Циркуляторный транспорт. Инсулин, поступая в кровь воротной вены, доставляется в печень и далее в системный кровоток. Инсулин не имеет белка-носителя в плазме (в отличие от инсулиноподобных факторов роста), и поэтому его не превышает 10 мин.
Физиологические эффекты инсулина (основные мишени - мышечная и жировая ткань, печень).
• Клеточные механизмы действия инсулина. Инсулин соединяется с инсулиновым рецептором (Ш.) плазмолеммы и активирует его тирозинкиназный участок, что приводит к аутофосфорилированию. Активация рецептора приводит к образованию в клетке мультиферментного комплекса, способного осуществить фосфорилирование и дефосфорилирование тирозиновых остатков в молекулах белков, что и определяет влияние инсулина на метаболические процессы.
В формировании фосфорилирующего эффекта участвует белок-посредник IRS-1, который обеспечивает проведение сигнала на различные молекулы, среди которых наиболее изучен фермент фосфатидилинозитол-3-киназа, которая стимулирует перемещение белка-переносчика глюкозы ГЛЮТ-4 из цитозольных везикул в плазматическую мембрану.
Дефосфорилирующий эффект осуществляется путем активирования инсулиновым рецептором тирозиновых протеинфосфатаз, одной из мишенью которых является IRS-белок, что приводит к его инактивации и, следовательно, ингибированию эффектов инсулина.
Влияние инсулина на обмен углеводов.
· Стимулирует транспорт глюкозы через мембраны мышечных и жировых клеток (на них увеличивается количество белков-переносчиков ГЛЮТ 4, образующих гидрофильные трансмембранные каналы для глюкозы, которые в 40 раз увеличивают транспорт глюкозы в клетки).
· Снижает уровень глюкозы в крови (гипогликемия).
· Усиливает синтез гликогена (активация гликогенсинтетазы).
· Активирует глюкокиназу (усиление использования глюкозы).
· Активирует глюкозо-6-фосфатдегидрогеназу (использование глюкозы в пентозофосфатном пути).
· Ингибирует глюкозо-6-фосфатазу в печени и почках (уменьшает выход глюкозы в кровь).
· Ингибирует глюконеогенез, снижая образование через геном ключевого фермента глюконеогенеза (фосфоэнолпируваткарбоксикиназу).
Влияние инсулина на липидный обмен.
· Тормозит липолиз и стимулирует липогенез (усиливает синтез жирных кислот и триглицеридов).
· Снижает кетогенез за счет стимуляции окисления кетоновых тел (ацетоацетата, (3-оксибутирата) в цикле трикарбоновых кислот.
На обмен белков инсулин оказывает анаболическое влияние.
· Усиливает транспорт аминокислот и синтез белков в мышечной и жировой ткани, в печени.
· Тормозит протеолиз и оказывает антикатаболическое действие.
Катаболизм инсулина происходит во многих тканях, но преимущественно в печени, почках и плаценте, в результате восстановления дисульфидных мостиков (фермент глютатион-инсулин-трансгидрогеназа) и гидролиза цепей (фермент инсулиназа).
Регуляция продукции инсулина.
Гуморальная регуляция. Повышение глюкозы в крови - основной стимулятор секреции инсулина (пороговая величина глюкозы - около 5 ммоль/л, максимальный ответ - при концентрации около 25 ммоль/л). На р-клетках имеются белок-транспортер глюкозы - ГЛЮТ 2. Стимулируют секрецию инсулина аминокислоты лейцин, аргинин, лизин.
Эндокринная регуляция.
Глюкагон, глюкокортикоиды, эстрогены, прогестерон и гастроинте-СТИНальные гормоны (особенно секретин, холецистокинин-панкреозимин) стимулируют секрецию инсулина. Последние подготавливают ин-сулиновую систему к предстоящему при пищеварении всасыванию глюкозы в кровь. Соматостатин и катехоламины ингибирует секрецию инсулина
Нервная регуляция (вегетативная нервная система).
Парасимпатические влияния стимулируют секрецию инсулина (ацетилхолин через М-холинорецепторы) и катехоламины.
Симпатические влияния тормозят секрецию инсулина через агадренорецепторы и стимулируют - через Рг-адренорецепторы (суммарный эффект - тормозящий).
Эндокринная функция а-клеток, вырабатывающих глюкагон (30 - 120 нг/л плазмы, период полураспада 5-10 мин).
Физиологические эффекты глюкагона.
Влияние на обмен углеводов и белков.
· Стимулирует продукцию глюкозы в печени в результате гликогенолиза (активирует фосфорилазу).
· Тормозит гликогенсинтетазу и синтез гликогена.
· Стимулирует катаболизм белков и глюконеогенез – образование глюкозы из аминокислот и лактата.
· Усиливает вход в печень аланина и образование мочевины.
Влияние на обмен липидов.
· Усиливает липолиз в жировой ткани (через аденилатциклазу и цАМФ).
· Увеличивает поставку в печень свободных жирных кислот.
· Усиливает сокращение сердца.
Влияние на секрецию других гормонов.
· Стимулирует клетки и секрецию инсулина.
· Стимулирует мозговой слой надпочечников и секрецию катехоламинов.
· Стимулирует секрецию кальцитонина в щитовидной железе.
Регуляция секреции глюкагона.
Секреция стимулируется гипогликемией, аминокислотой аргинином, свободными жирными кислотами, холецистокинин-панкреозимином, симпатической нервной системой. Секреция тормозится гипергликемией, соматостатином, секретином.
Эндокринная функциия δ –клеток, вырабатывающих соматостатин.
Эффекты соматостатина.
· Тормозит секрецию инсулина и глюкагона.
· Снижает всасывание глюкозы в кишечнике.
· Тормозит секрецию соляной кислоты, пепсинов и моторику в желудке, секрецию ферментов поджелудочной железы.
Регуляция секреции: стимуляторами секреции служат глюкагон и все активаторы секреции инсулина.
Тема: Эндокринная система
Цель: а) знать структурно-функциональную организацию эндокринной системы, образование, выделение, перенос и распад гормонов, их основные механизмы действия, саморегуляцию эндокринной системы, функциональные связи желез внутренней секреции и нервной системы, нейросекреты гипоталамуса; основные виды влияний гормонов на органы и системы организма; методы исследования эндокринной функции;
б) уметь оценивать роль эндокринных механизмов в регуляции гомеостаза, адаптации организма к условиям внешней среды, в процессах высшей нервной деятельности и поведения.
Мотивация темы: гормональная регуляция является обязательным элементом поддержания гомеостаза, участвует в реализации высших психических функций: мотивации, эмоции, сна, памяти. Адаптивные реакции организма, в том числе стресс, протекают при обязательном участии гормонов. Нарушения функции эндокринных желез являются одной из наиболее распространенных форм патологии, с которой сталкивается врач любой специальности. Знания в области эндокринологии необходимы как для своевременной и точной диагностики нарушений, так и для корректного применения гормонов в лечебной практике.
САМОСТОЯТЕЛЬНАЯ РАБОТА СТУДЕНТОВ ВО ВНЕАУДИТОРНОЕ ВРЕМЯ
Задание 1. Изучить теоретический материал занятия, используя следующую логическую структуру учебного материала.
Эндокринная система - совокупность взаимосвязанных и различным образом организованных эндокринных клеток, осуществляющая централизованное управление функций организма с помощью сигнальных молекул - гормонов. Гормоны - внеклеточные сигнальные регуляторы, образующиеся специализированными эндокринными клетками и обладающие дистантным действием..
Виды объединения эндокринных клеток.
- Формируют эндокринные железы.
- Образуют часть железистого (например, островки поджелудочной железы, гонады) или нежелезистого (например, почки, сердце) органа.
- Эндокринные клетки диффузно расположены в органах (гастроинтестинальная система).
- Образование эндокринной системы как результат взаимодействия эндокринных структур.
Виды действия гормонов.
Эндокринное (дистантное) действие - гормоны эндокринной железы действуют на клетки-мишени других органов и тканей.
Паракринное действие - гормоны эндокринной клетки действуют на соседние клетки в пределах одного органа (например, гастроинтестинальные гормоны, простагландины).
Аутокринное действие - гормоны эндокринной клетки действуют на эту же клетку (например, простагландины).
Химическая характеристика гормонов.
Основные классы гормонов.
1.Белково-пептидные гормоны (например, гормоны гипофиза).
2. Производные аминокислот (из тирозина образуются адреналин, норадреналин, дофамин, тироидные гормоны, из триптофана - мелатонин, серотонин).
3. Стероидные гормоны (кортикостероиды, половые гормоны, 1,25-(ОН)2-витамин D3).
Основные фрагменты молекулы гормонов.
Адресный фрагмент обеспечивает связывание гормона с циторецептором например, α -субъединица лютеинизирующего гормона. (Снижение этого сродства приводит к гормонорезистентности.) Актон обеспечивает специфичность действия гормона (например,β - субъединица лютеинизирующего гормона). Вспомогательные фрагменты служат для изменения его действия, определяют его иммунологические свойства.
Секреция гормонов.
Экзоцитоз секреторных гранул (пептидные гормоны, катехоламины) стимулируется Са2+, поступившим в цитозоль через Са2+-каналы при деполяризации мембраны клетки или (и) из гладкой ЭПС при действии инозитол-трифосфата.
Освобождение гормона из белковосвязанной формы (тироксин из иодированного тироглобулина).
Свободная диффузия гормонов, растворимых в липидах мембраны (например, кортикостероиды, половые гормоны).
Циркуляторный транспорт гормонов (основные формы их транспорта в крови).
Свободная, несвязанная форма составляет от сотых долей процента (например, Т4 и Т3) до 100 % (например, инсулин). Только эта форма гормона действует на клетки-мишени.
Специфическое связывание с глобулинами (70 - 80 %) обеспечивает создание запаса и связывание избытка гормона в крови, защиту его от инактивации и от потери через почки. (Виды глобулинов - транскортин, тироксин-связывающий глобулин, прогестин, тестостерон-эстрогенсвязывающий глобулин.)
♦ Неспецифическое связывание с альбуминами крови (5 - 10 %). Благодаря низкой прочности связи они являются основным источником свободных гормонов в крови. (Небольшая часть гормона связана с форменными элементами крови.)
♦ Периферическая конверсия гормонов - превращение гормонов в органах и тканях, отдаленных от эндокринной железы, в результате которого изменяется активность гормона (например, превращение тироксина в трииодтиронин в печени, тестостерона в дигидротестостерон - в простате).
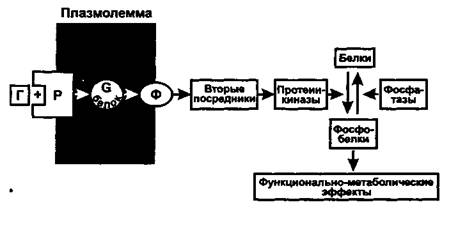
Рис. 1. Общая схема действия гормонов через вторые посредники. Г - гормон, Р - рецептор, Ф - фермент
Физиологические эффекты гормонов.
Клетки - (ткани-, органы-) мишени, избирательно реагирующие на гормоны, имеют циторецепторы - белковые приемники и преобразователи гормонального сигнала. (Есть данные, что на один рецептор может действовать более, чем один гормон.) Циторецепторы расположены на клеточной мембране или в ядре (500-30000 рецепторов в клетке). Максимальный эффект гормона наблюдается при его концентрации, обеспечивающей занятость всего 5 - 10 % общего числа рецепторов (феномен «резервных» рецепторов). Изменение числа рецепторов является способом регуляции гормонального ответа (активация и ингибирование синтеза рецепторов, интернализация рецепторов путем эндоцитоза).Гормоны, плохо проникающие в клетку (белково-пептидные, катехоламины), соединяются с рецепторами на клеточной мембране и вызывают внутриклеточные эффекты через вторые посредники. Большинство гормонов после присоединения к рецепторам активируют регулируемый гуанилнуклеотидами С-белок мембраны. Имеется несколько видов G-белков (Gs, Gi, Gq, G0), которые влияют на ферменты (например, аденилатциклазу, фосфолипазу С), образующие различные вторые посредники (например, цАМФ, инозитолтрифосфат, диацилглицерол). При этом G-белки могут как активировать (Gs), так и ингибировать (GO образование вторых посредников. Вторые посредники, в свою очередь, активируют соответствующие им протеинкиназы, фосфорилирующие различные клеточные белки по остаткам серина и треонина, что изменяет их функциональную активность (чаще увеличивает). Наряду с этим активированные G-белки оказывают выраженное влияние на проницаемость кальциевых и калиевых каналов мембраны.В действии гормонов, плохо проникающих в клетки, можно выделить 4 системы: аденилатциклазную, фосфоинозитидную, гуанилатциклазную и тирозинкиназную.
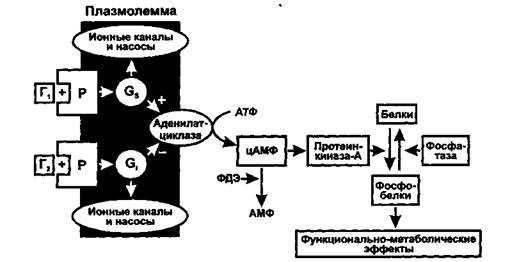
Рис. 2 Схема действия гормонов через аденилатциклазную систему. Г - гормон, Р - рецептор, Ф - фермент, ФДЭ - фосфодиэстераза.
• Аденилатциклазная система. Эту систему используют гормоны, осуществляющие свой эффект через аденилатциклазу и синтез второго посредника циклического АМФ (3,5-цАМФ). Действие гормона может как активировать, так и тормозить его образование.
Комплекс «гормон + рецептор» активирует Gs-белок, который активирует аденилатциклазу и образование цАМФ (например, так действуют адреналин через рУр2-рецепторы, АДГ через У2-рецепторы, глюкагон, АКТГ, ТТГ, ФСГ, ЛГ, МСГ, кортиколиберин, соматолиберин, кальцитонин, хориогона-дотропин человека, паратгормон и др.).
Комплекс «гормон + рецептор» активирует Gi-белок, который тормозит аденилатциклазу и образование цАМФ (например, так действуют адреналин через а2-рецепторы, ангиотензин II через АТ-рецепторы, аденозин через Ai-рецепторы, соматостатин, опиоиды через, 5- и к-рецепторы, ацетилхолин через Мг-холинорецепторы гладкомышечных клеток).
цАМФ влияет на функции клетки через изменение активности протеинфосфокиназы-А, фосфорилирующей различные клеточные белки: мембранные (изменяется ее проницаемость), ферменты (изменяется метаболизм клетки), ядерные (изменение транскрипции), рибосом (изменение трансляции), микротрубочек (двигательные эффекты, изменение формы клетки) и др.
• Фосфоинозитидная система. Эту систему используют гормоны, которые, действуя через Оч-белок, активируют в мембране фосфолипазу С. Она образует из фосфолипида мембраны (фосфатидилинози-толдифосфата) два посредника - инозитол-1,4,5-трифосфат (ИФз) и 1,2- диацилглицерол (ДАГ).
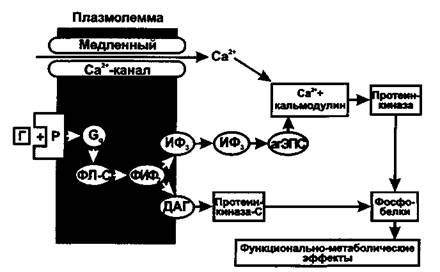
Рис. 3. Схема действия гормонов через фосфоинозитидную систему. Г - гормон, Р - рецептор, ФЛ-С - фосфолипаза С, ФИФ2 - фосфатиди-линозитолдифосфат, ИФ3 - инозитолтрифосфат, ДАГ - диацилглицерол
ИФз, переходя в цитоплазму, действует на рецепторы гладкой ЭПС, что приводит к открыванию Са2+-каналов и выходу Са2+ в цитозоль. Са2+ при этом выступает в качестве третьего посредника, который связывается с белком кальмодулином и активирует Са2+-кальмоду-линзависимую протеинкиназу, фосфорилирующую эффекторные белки, влияющие на функции клеток, их пролиферацию и дифференцировку (регулирует активность аденилатциклазы и синтез цАМФ, фосфолипазы-Аг и синтез простагландинов, тромбоксана, простациклина. гуанилатциклазы и синтез цГМФ, фосфодиэстеразы и разрушение цАМФ и цГМФ; участвует в регуляции секреции инсулина, гормонов щитовидной железы, гипофиза и надпочечников, кишечной секреции и секреции нейромедиаторов). (Роль Са2+ как посредника действия гормонов обусловлена его сильным влиянием при низких концентрациях на метаболизм и функции клетки. Гормоны могут увеличивать концентрацию Са2+ в цитозоле как в результате открывания Са2+-каналов клеточной мембраны, так выхода Са2+ из гладкой ЭПС. В клетке есть специальные белки, связывающие Са2+ при низкой концентрации -кальмодулин, гуанилатциклаза, тропонин С и др. Присоединение Са2+ к этим белкам изменяет их конфигурацию и свойства. Это приводит к изменению многочисленных функций клеток - метаболической, секреторной, сократительной, биоэлектрической и др.
ДАГ, оставаясь на внутренней поверхности мембраны, активирует протеинкиназу С, фосфорилирующую эффекторные белки (например, активирует киназу легкой цепи миозина), а также синтез простагландинов из арахидоновой кислоты, что изменяет функции клетки.
Через ИФз и ДАГ действуют адреналин (oci -рецепторы), вазопрессин (Vi), ацетилхолин, ангиотензин П, гистамин, холецистокинин, окситоцин, ВИП и др.
• Гуанилатциклазная система. Некоторые гормоны (например, ацетилхолин, цитокины) действуют через гуанилатциклазу, образующую второй и посредник - циклический ГМФ (цГМФ). Гуанилатциклаза существует в двух формах - мембраносвязанной и растворимой в цитозоле. Мембраносвязанная гуанилатциклаза является частью рецептора плазмолеммы, ее активация гормоном (например, На+- уретическим) увеличивает образования цГМФ, который стимулирует протеинкиназу G, фосфорилирующую белки. Растворимая в цитозоле гуанилатциклаза содержит в своем составе гем, с которым связывается сигнальная молекула - NО (оксид азота образуется из аргинина в ряде клеток - эндотелии, нейронах и др. под действием ацетилхолина, механических раздражителей и др.). Активированная оксидом азота гуанилатциклаза через протеинкиназу G фосфорилирует белки (например, так действует ацетилхолин, вызывая расширение сосудов).
Тирозинкиназная система характеризуется особыми рецепторами, которые путем фосфорилирования и дефосфорилирования сигнальных белков по тирозиновым остаткам, регулируют процессы роста, дифференцировки, выживания, метаболизма и смерти клеток. Эту систему используют некоторые гормоны - инсулин, инсулиноподобные факторы роста (или соматомедины), факторы роста эпидермиса, тромбоцитов и др. Многие рецепторы к таким гормонам содержит в своем составе тирозинкиназу, которая при действии гормона аутофосфорилирует внутреннюю часть рецептора по остаткам тирозина. Активированный таким образом рецептор передает через сложный и недостаточно изученный комплекс посредников фосфорилирующее и дефосфорилирующее влияние по остаткам тирозина на многие сигнальные молекулы цитоплазмы и ядра. Фосфорилирование и дефосфорилирование белков может приводить как к их активации, так и инактивации.
В действительности гормоны оказывают преимущественно разветвленные эффекты. Это связано с тем, что комплекс Г+Р действует с разной степенью на несколько G-белков. В свою очередь активированные G-белки действуют на несколько эффекторов (ферментов, каналов).
Клеточные эффекты липорастворимых гормонов, легко проникающих через клеточные мембраны (стероидные и тироидные гормоны, 1,25-(ОН)г-витамин D3). Гормон соединяется с ядерным рецептором. Рецепторы представляют собой белки (факторы транскрипции), прочно связанные с ДНК хроматина и дополнительными ядерными белками. Каждый рецептор содержит несколько специфических участков (доменов): ДНК-связывающий домен, гормонсвязывающий домен (гидрофобный «карман»), домен, активирующий транскрипцию и др. Активируются процессы транскрипции (образование мРНК с помощью фермента РНК-полимеразы II) и трансляции (взаимодействие рРНК, тРНК и мРНК, индуцирующее синтез белков и дифференцировку клеток). Например, глюкокортикоиды стимулируют гены, обеспечивающие синтез.
Парагипофизарный путь.
Прямой нервный контроль секреции гормонов осуществляется в мозговом слое надпочечников, эпифизе, гипоталамусе (его нейросекреторных ядрах); в этих железах происходит преобразование нервного импульса в нейросекреторный процесс.
Эндокринная (негипофизарная) регуляция осуществляется в результате влияния гормонов периферических эндокринных желез друг на друга (например, торможение кортизолом секреции тироксина, соматостатином секреции инсулина и глюкагона).
Гуморальная регуляция секреции инсулина, альдостерона, паратгормона осуществляется соответственно уровнем глюкозы, Na+ и Са2+ в крови.
Методы исследования желез внутренней секреции.
- Удаление железы или подавление ее активности.
- Пересадка железы или введение гормонов.
- Определение количества гормонов и их метаболитов в биологических жидкостях (крови, моче, ликворе) биохимическими, иммунорадиологическими и имму-нофлюоресцентными методами анализа.
- Функциональные нагрузочные пробы (например, тест на толерантность к глюкозе).
Физиология гипофиза.
Гормоны нейрогипофиза.
Антидиуретический гормон (АДГ) или вазопрессин (нонапептид супраоптических и паравентрикулярных ядер гипоталамуса, концентрация в плазме 1-13 нг/л).
Эффекты АДГ (вазопрессина).
Увеличивает реабсорбцию воды в дистальных канальцах и собирательных трубочках почек, что приводит к уменьшению диуреза (эффект реализуется через У2-рецепторы, повышение уровня цАМФ и увеличение числа водных каналов в клеточной мембране).
В более высоких концентрациях усиливает сокращения гладких мышц и вызывает сужение сосудов.
Активирует центр жажды и питьевое поведение (одновременно тормозит секрецию ренина).
Участвует в механизмах формирования памяти (стимулирует обучение).
Регуляция секреции АДГ.
Главным регулятором секреции АДГ является осмоляльность плазмы, которая контролируется осморецепторами гипоталамуса, сосудов, тканей. Гиперосмия (>290 мосм/кг) стимулирует секрецию АДГ, гипоосмия (< 280 мосм/кг) тормозит секрецию АДГ.
Второй регулятор секреции АДГ - колебания объема циркулирующей крови (ОЦК) и АД, которые регистрируются барорецепторами аорты, сонной артерии, легочной артерии, волюморецепторами предсердий и вен. Гиповолемия (снижение ОЦК более чем на 5 %) и снижение АД стимулируют секрецию, гиперволемия (увеличение ОЦК) и повышение АД тормозят секрецию АДГ.
Другие факторы, влияющие на секрецию АДГ.
Стимулируют секрецию АДГ симпатоадреналовая система (через Р-адрено-рецепторы), ангиотензин II, гистамин, ацетилхолин.
Тормозят секрецию АДГ натрийуретический гормон предсердий, этанол, переохлаждение.
Окситоцин (образуется в паравентрикулярных и супраоптических ядрах гипоталамуса, концентрация в плазме 3,2 мМЕ/л).
Эффекты окситоцина реализуются через поступление Са +:
Усиливает сократительную активность матки во время родов (эстрогены увеличивают чувствительность матки к окситоцину, а прогестерон снижает).
Усиливает сокращение гладкой мускулатуры протоков молочных желез и выделение молока.
Стимулирует продукцию простагландинов в эндометрии, которые совместно с окситоцином вызывают начало родов (мобилизуют Са+).
Участвует в механизмах формирования памяти (нарушает запоминание).
Регуляция секреции окситоцина: рефлекторная стимуляция секреции с
рецепторов растяжения шейки матки и влагалища во время родов, сосков молочных желез при кормлении грудью; условнорефлекторная – на крик ребенка и кормление.
Околощитовидные железы.
Физиологические эффекты паратгормона - повышает уровень Са2+ в крови в результате действия следующих механизмов.
· Повышение активности остеоцитов и остеокластов (усиление резорбции костной ткани).
· Увеличение реабсорбции Са2+ в дистальных почечных канальцах и снижения реабсорбции фосфатов в проксимальных почечных канальцах (фосфатурия).
· Усиление всасывания Са2+ в тонком кишечнике (действует совместно с витамином D3).
Эффект паратгормона связан со стимуляцией аденилатциклазы, увеличением уровня цАМФ и активности протеинфосфокиназы А в клетках-мишенях.
Регуляция продукции паратгормона (суточный ритм секреции: в ночные часы в 2 - 3раза больше, чем в дневное время). Основной стимулятор секреции - гипокальциемия (ускоряет трансформацию пропаратгормона в активный гормон). Ингибитор секреции - снижение уровня Mg2+ в крови.
Эндокринные функции островков Лангерганса поджелудочной железы: α-клетки
(25 %) продуцируют глюкагон; Р-клетки (60 %) - инсулин; β-клетки (10 %) - соматостатин (все поли пептиды).
Эндокринные функции β-клеток, вырабатывающих инсулин (в крови находится в
свободной форме, концентрация в плазме 6-24 мМе/л).
Секреция инсулина стимулируется глюкозой крови, которая проникает в β-клетки преимущественно с помощью транспортера ГЛЮТ 2, что приводит к повышению уровня АТФ и закрытию АТФ-зависимьгх К+ - каналов. Возникает деполяризация мембраны до КУД и возникникает ПД. Вход Са2+ в клетку во время ПД запускает экзоцитоз гранул инсулина и открытие К+-каналов. Секреция инсулина осуществляется в кровь панкреатической вены.
Циркуляторный транспорт. Инсулин, поступая в кровь воротной вены, доставляется в печень и далее в системный кровоток. Инсулин не имеет белка-носителя в плазме (в отличие от инсулиноподобных факторов роста), и поэтому его не превышает 10 мин.
Физиологические эффекты инсулина (основные мишени - мышечная и жировая ткань, печень).
• Клеточные механизмы действия инсулина. Инсулин соединяется с инсулиновым рецептором (Ш.) плазмолеммы и активирует его тирозинкиназный участок, что приводит к аутофосфорилированию. Активация рецептора приводит к образованию в клетке мультиферментного комплекса, способного осуществить фосфорилирование и дефосфорилирование тирозиновых остатков в молекулах белков, что и определяет влияние инсулина на метаболические процессы.
В формировании фосфорилирующего эффекта участвует белок-посредник IRS-1, который обеспечивает проведение сигнала на различные молекулы, среди которых наиболее изучен фермент фосфатидилинозитол-3-киназа, которая стимулирует перемещение белка-переносчика глюкозы ГЛЮТ-4 из цитозольных везикул в плазматическую мембрану.
Дефосфорилирующий эффект осуществляется путем активирования инсулиновым рецептором тирозиновых протеинфосфатаз, одной из мишенью которых является IRS-белок, что приводит к его инактивации и, следовательно, ингибированию эффектов инсулина.
Влияние инсулина на обмен углеводов.
· Стимулирует транспорт глюкозы через мембраны мышечных и жировых клеток (на них увеличивается количество белков-переносчиков ГЛЮТ 4, образующих гидрофильные трансмембранные каналы для глюкозы, которые в 40 раз увеличивают транспорт глюкозы в клетки).
· Снижает уровень глюкозы в крови (гипогликемия).
· Усиливает синтез гликогена (активация гликогенсинтетазы).
· Активирует глюкокиназу (усиление использования глюкозы).
· Активирует глюкозо-6-фосфатдегидрогеназу (использование глюкозы в пентозофосфатном пути).
· Ингибирует глюкозо-6-фосфатазу в печени и почках (уменьшает выход глюкозы в кровь).
· Ингибирует глюконеогенез, снижая образование через геном ключевого фермента глюконеогенеза (фосфоэнолпируваткарбоксикиназу).
Влияние инсулина на липидный обмен.
· Тормозит липолиз и стимулирует липогенез (усиливает синтез жирных кислот и триглицеридов).
· Снижает кетогенез за счет стимуляции окисления кетоновых тел (ацетоацетата, (3-оксибутирата) в цикле трикарбоновых кислот.
На обмен белков инсулин оказывает анаболическое влияние.
· Усиливает транспорт аминокислот и синтез белков в мышечной и жировой ткани, в печени.
· Тормозит протеолиз и оказывает антикатаболическое действие.
Катаболизм инсулина происходит во многих тканях, но преимущественно в печени, почках и плаценте, в результате восстановления дисульфидных мостиков (фермент глютатион-инсулин-трансгидрогеназа) и гидролиза цепей (фермент инсулиназа).
Регуляция продукции инсулина.
Гуморальная регуляция. Повышение глюкозы в крови - основной стимулятор секреции инсулина (пороговая величина глюкозы - около 5 ммоль/л, максимальный ответ - при концентрации около 25 ммоль/л). На р-клетках имеются белок-транспортер глюкозы - ГЛЮТ 2. Стимулируют секрецию инсулина аминокислоты лейцин, аргинин, лизин.
Эндокринная регуляция.
Глюкагон, глюкокортикоиды, эстрогены, прогестерон и гастроинте-СТИНальные гормоны (особенно секретин, холецистокинин-панкреозимин) стимулируют секрецию инсулина. Последние подготавливают ин-сулиновую систему к предстоящему при пищеварении всасыванию глюкозы в кровь. Соматостатин и катехоламины ингибирует секрецию инсулина
Нервная регуляция (вегетативная нервная система).
Парасимпатические влияния стимулируют секрецию инсулина (ацетилхолин через М-холинорецепторы) и катехоламины.
Симпатические влияния тормозят секрецию инсулина через агадр






