(чувство осязания, прикосновения).
Тактильная чувствительность от кожи затылка, выпуклой части ушной раковины, шеи, туловища и конечностей проводится по спинномозговым нервам, а от кожи, лица, вогнутой стороны ушной раковины, передних отделов волосистой части головы – по тройничному нерву и лишь от кожи задней стенки наружного слухового прохода – по блуждающему нерву.
Тела первых нейронов, воспринимающих тактильную чувствительность из кожи затылка, выпуклой части ушной раковины, шеи, туловища и конечностей, заложены в спинномозговых узлах. Их периферические отростки (дендриты) идут в составе ветвей спинномозговых нервов к коже, заканчиваясь в ней рецепторами (тельца Мейснера). Центральные отростки (аксоны) в составе задних корешков вступают в спинной мозг и большей своей частью контактируют с клетками желатинозной субстанции. Меньшая же часть волокон, не заходя в серое вещество заднего рога, следует в составе пучков Голля и Бурдаха в продолговатый мозг, где вступают в контакт с клетками ядер этих пучков. Клетки желатинозной субстанции и ядер пучков Голля и Бурдаха являются вторыми нейронами.
Аксоны вторых нейронов, заложенных в желатинозной субстанции, проходят через переднюю белую спайку спинного мозга на противоположную сторону и в передне-наружном отделе бокового канатика образуют передний спинно-таламический тракт. Далее эти волокна вместе с волокнами бокового спинно-таламического тракта в верхнем отделе моста присоединяются к медиальной петле и заканчиваются в латеральном ядре зрительного бугра, где заложены тела третьих нейронов.
Аксоны вторых нейронов, лежащих в ядрах нежного и клиновидного пучков, также переходят на противоположную сторону и в составе медиальной петли подходят к латеральному ядру зрительного бугра, вступая в синаптическую связь с телами третьих нейронов. Аксоны третьих нейронов в составе таламокортикального тракта проходят через заднюю треть заднего бедра внутренней капсулы к коре задней центральной извилины и верхней теменной дольки, где заложено ядро кожного анализатора.
Поскольку в спинном мозге одна часть волокон, проводящих тактильную чувствительность, поднимается по своей стороне, а другая - по противоположной, то при одностороннем поражении последнего расстройства чувствительности может и не отмечаться.
Первые нейроны, воспринимающие тактильную чувствительность от кожи лица, передних отделов волосистой части головы и наружного уха, заложены в узлах тройничного и блуждающего (верхний узел) нервов. Периферические их отростки в составе нервов следуют к коже, заканчиваясь в ней рецепторами, а центральные отростки (аксоны) подходят к указанным выше чувствительным ядрам этих нервов. Здесь заложены тела вторых нейронов, аксоны которых переходят на противоположную сторону и в составе медиальной петли идут к латеральному ядру зрительного бугра, где вступают в контакт с телами третьих нейронов. Аксоны последних также достигают коры задней центральной извилины и верхней теменной дольки.
Проводящий путь пространственной кожной чувствительности- стереогноза (узнавание предметов на ощупь)
Путь этого вида кожной чувствительности аналогичен пути той части тактильной чувствительности, которая проводится по пучкам задних канатиков. Первыми нейронами являются ложно-униполярные клетки спинальных ганглиев. Дендриты их заканчиваются в коже рецепторами, а аксоны в составе задних корешков вступают в спинной мозг и, минуя задние рога, поднимаются в задних канатиках к ядрам нежного и клиновидного пучков продолговатого мозга. Здесь лежат тела II нейронов. Аксоны их, перейдя на противоположную сторону, в составе медиальной петли достигают латерального ядра зрительного бугра и контактируют с телами III нейронов. Аксоны III нейронов следуют через заднюю треть заднего бедра внутренней капсулы в составе таламокортикального тракта в кору верхней теменной дольки.
Корковый конец кожного анализатора, получив и переработав разнообразную информацию из внешней среды, передает её исполнительной системе, представленной пирамидным и экстрапирамидными путями. По ним осуществляется ответная реакция на воспринятые раздражения.
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Отличительной особенностью этого анализатора является вынесение из мозга на периферию цепи синаптически связанных друг с другом трех нейронов, заложенных в сетчатой оболочке глазного яблока. Первыми нейронами являются зрительные фоторецепторные клетки, тела которых расположены в наружном зернистом (ядерном) слое сетчатки. Периферические отростки этих клеток заканчиваются рецепторами — палочками или колбочками, погруженными в пигментный слой сетчатой оболочки.
Количество палочек в сетчатке достигает 125 – 130 млн., а колбочек -5 – 8 млн. Плотность расположения их в сетчатке не одинакова. В области центральной ямки желтого пятна располагаются только колбочки. К периферии от желтого пятна количество их быстро уменьшается, а количество палочек увеличивается. Колбочки обеспечивают дневное зрение (острота зрения и цветоощущение), а палочки — ночное или сумеречное зрение (светоощущение, световая и темновая адаптация). Световые раздражения преобразуются палочками и колбочками в нервные импульсы, которые фоторецепторными клетками передаются вторым нейронам — биполярным клеткам, расположенным во внутреннем зернистом слое сетчатки.
С одной биполярной клеткой контактируют от 1 до 30 колбочек или до 500 палочек. В области центральной ямки каждая биполярная клетка контактирует лишь с одной колбочкой. От биполярных клеток (вторых нейронов) нервные импульсы передаются третьим нейронам — мультиполярным ганглиозным клеткам, расположенным в ганглиозном слое сетчатки. Аксоны этих клеток, сближаясь, образуют зрительный нерв. Место схождения волокон носит название диска зрительного нерва. Его диаметр в норме составляет от 1 до 2 мм. В этом месте сетчатки нет ни колбочек, ни палочек, отчего оно получает название слепого пятна. Зрительные нервы через зрительные каналы покидают глазницы и, войдя в полость черепа, в области диафрагмы турецкого седла, между дном 3 желудочка и гипофизом образуют частичный перекрест, причем перекрещиваются волокна только от медиальных (носовых) половин сетчатых оболочек обоих глаз (рис. 19). Волокна же от латеральных (височных) половин сетчаток не перекрещиваются. От перекреста начинаются зрительные тракты, содержащие волокна от одноименных половин сетчатки обоих глаз. Так, правый зрительный тракт содержит неперекрещенные волокна правой (височной) половины сетчатки правого глаза и перекрещенные волокна от правой (носовой) половины сетчатки левого глаза. Соответственно, в левом зрительном тракте проходят неперекрещенные волокна от левой (височной) половины сетчатки левого глаза и перекрещенные волокна от левой (носовой) половины правого глаза. Зрительные тракты, огибая ножки мозга, подходят к подкорковым центрам зрения. Большая часть их волокон оканчивается в латеральных коленчатых телах, а меньшая — в подушках зрительных бугров и верхних бугорках четверохолмия, вступая в контакт с телами четвертых нейронов.
Часть аксонов ганглиозных клеток сетчатой оболочки (третьих нейронов), минуя подкорковые центры зрения, следуют в кору затылочной доли к ядру зрительного анализатора. В результате ускоряется поступление зрительной информации к корковому концу зрительного анализатора и корковые нейроны подготавливаются к восприятию импульсов, идущих по переключательным путям, т. е. через подкорковые центры. Соответственно укорачивается и время реакции коры на зрительное раздражение. Кроме того, часть аксонов ганглиозных клеток сетчатки следует к корковым концам двигательного и слухового анализаторов, обеспечивая межанализаторную интеграцию. Необходимо также отметить, что в составе зрительных трактов выявлены нервные волокна, которые следуют к подкорковым центрам слухового анализатора (медиальным коленчатым телам и нижним бугоркам четверохолмия), ядрам гипоталамуса, ретикулярной формации, ядрам глазодвигательных нервов своей и противоположной стороны и гиппокампу, осуществляя прямую связь зрительного анализатора с филогенетически более старыми структурами мозга. Это говорит об участии последних в анализе зрительной сигнализации.
Аксоны клеток латерального коленчатого тела и подушки зрительного бугра (4 нейронов) проходят через задний отдел заднего бедра внутренней капсулы и, образуя зрительную лучистость, следуют к ядру зрительного анализатора, заложенному в области шпорной борозды (поле 17), а также к 18 и 19 корковым полям затылочной доли (рис. 20). На поле 17 проецируются все слои латерального коленчатого тела, причём конвергенция импульсов со своей и противоположной стороны совершается в четвертом гранулярном слое коры, благодаря чему создаются условия для бинокулярного стереоскопического зрения, требующего сочетанных движений обоих глаз. Поэтому от поля 17 начинается эфферентный путь, направляющийся к центрам конвергенции и дивергенции, расположенным, по-видимому, в области ядер глазодвигательного нерва.
Из подушки зрительного бугра волокна следуют к 18 и 19 корковым полям. Поле 18 получает также информацию от поля 17 посредством коротких ассоциативных волокон и от глазодвигательного поля 8 по лобно-затылочному продольному пучку. Благодаря этому осуществляется зрительное восприятие образов. Эфферентные волокна
| Рис. 19.Пути зрительного анализатора
| |








поля 18 идут к верхним бугоркам четверохолмия, которые управляют согласованными движениями глаз. Поле 19 посредством ассоциативных волокон связано с полем 18 и, по-видимому, осуществляет высшие зрительные функции — узнавание зрительных образов.
Необходимо подчеркнуть, что в составе периферического и центрального звеньев зрительного анализатора содержатся не только афферентные, но и эфферентные волокна. Последние, возникнув в коре, следуют к латеральному коленчатому телу, подушке зрительного бугра, задним бугоркам четверохолмия, а также к сетчатке глаза, оказывая на них корригирующее влияние.
Как отмечалось выше, часть волокон зрительного тракта заканчивается в верхних бугорках четверохолмия синапсами с телами четвертых нейронов. Часть аксонов 4 нейронов переключается на клетки глубокого слоя верхних бугорков, аксоны которых переходят на противоположную сторону (дорзальный перекрест покрышки) и под названием текто-бульбарного и тектоспинального путей следуют к двигательным ядрам черепных нервов и передних рогов спинного мозга. По этому пути осуществляются рефлекторные ответные реакции на неожиданные световые раздражения (сторожевой рефлекс).


Рис. 20 Цитоархитектонические поля коркового конца зрительного анализатора (Московский институт мозга)
Другая часть аксонов 4 нейронов переключается на вегетативные ядра глазодвигательных нервов (ядра Якубовича) своей и противоположной стороны и на непарное срединное ядро. К этим ядрам подходят и прямые волокна из сетчатой оболочки глаза. Аксоны клеток вегетативных ядер покидают средний мозг в составе глазодвигательных нервов и подходят к ресничному узлу, где переключаются на следующий нейрон. Аксоны последних нейронов (клеток ресничного узла) в составе коротких ресничных нервов следуют к мышце, суживающей зрачок, и ресничной мышце. Через этот путь осуществляется зрачковый (пупиллярный) рефлекс, выражающийся в сужении зрачка при попадании в глаз пучка света, а также изменение кривизны хрусталика (аккомодация).
К эфферентным путям зрительного анализатора, помимо волокон, идущих из коры к подкорковым центрам и сетчатке, оказывающих на них корригирующее влияние, следует отнести пирамидный и экстрапирамидные пути, по которым корковый конец анализатора, переработав полученную информацию, посылает приказы к исполнительным органам—мышцам. Так осуществляется ответная двигательная реакция на воспринятые световые раздражения. Оптически деятельная часть сетчатой оболочки, выстилающая внутреннюю поверхность глазного яблока до плоской части ресничного тела или до зубчатой линии, позволяет каждому глазу охватить зрительным восприятием достаточно большое поле из окружающего нас мира. Оба глаза, при фиксации их на каком-либо предмете, по горизонтальному меридиану охватывают 180°, а по вертикальному- 120-130°. При этом поле зрения каждого глаза имеет определенные границы.
Изменения поля зрения при различных заболеваниях могут проявляться в виде сужения его границ, или в виде выпадения в нем отдельных участков и по характеру ограничения поля зрения можно установить локализацию поражения в том или ином отделе зрительного анализатора. Так, выпадение отдельного участка поля зрения указывает на частичное поражение зрительного нерва или сетчатой оболочки. Слепота одного глаза говорит о полном поражении зрительного нерва. При поражении хиазмы чаще всего страдают ее внутренние отделы, т. е. перекрестные волокна, идущие от обеих носовых половин сетчатой оболочки, что ведет к выпадению височных полей зрения обоих глаз. Полное разрушение хиазмы ведет к слепоте обоих глаз.
При поражении зрительного тракта или коркового ядра анализатора той же стороны наступает половинная слепота обоих глаз (гемианопсия). Так, при поражении зрительного тракта или коркового ядра анализатора слева на левом глазу выпадают не перекрещенные волокна, обслуживающие височную половину сетчатой оболочки, т. е. носовую половину поля зрения, а на правом перекрещенные волокна, обслуживающие носовую половину сетчатки, т. е. височную правую половину поля зрения. Иначе говоря, при наличии очага поражения слева выпадают правые половины полей зрения обоих глаз, а справа—левые.
СЛУХОВОЙ АНАЛИЗАТОР
Периферическая часть слухового анализатора представлена звукопроводящим (наружное и среднее ухо, пери- и эндолимфа внутреннего уха) и звуковоспринимающим аппаратом (кортиев орган).
Афферентный отдел (кондуктор) состоит из четырех нейронов (рис. 21). Первыми (рецепторными) нейронами являются биполярные клетки спирального узла. расположенного в одноименном канале основания спиральной пластинки улитки. Их периферические отростки (дендриты) подходят к чувствительным волосковым клеткам кортиева органа, являющимися рецепторами слухового анализатора. Центральные отростки (аксоны) биполярных клеток образуют улитковый нерв, который покидает внутреннее ухо через внутренний слуховой проход и вступает в мозг в области мосто-мозжечкового угла, заканчиваясь в вентральном и дорзальном слуховых ядрах. Некоторая часть аксонов биполярных клеток следует без перерыва, в кору верхней височной извилины. Эти прямые волокна создают преднастройку клеток коры для восприятия информации, поступающей по переключательным путям. Аксоны клеток вентральных ядер улитковых нервов следуют к ядрам верхней оливы (дорзальные ядра трапециевидного тела) и собственным ядрам трапециевидного тела своей и противоположной стороны. В этих ядрах заложены тела III нейронов. Большая часть аксонов клеток верхних олив и собственных ядер трапециевидного тела переходят на противоположную сторону, принимая участие в образовании трапециевидного тела. Последнее, таким образом, формируется аксонами клеток вентральных ядер (II нейроны), ядер верхних олив и собственных ядер трапециевидного тела (III нейроны) своей и противоположной стороны и представляет довольно толстый слой поперечных волокон, делящий мост на вентральную (основание) и дорзальную (покрышку) части.

| Gangl. spirale
(I нейрон)
| |


| Nucl.dorsalis n.cochlearis
(II нейрон)
| |
| Nucl.dorsalis corporis trapezoidei
(III нейрон)
| |
| Nucl.ventralis n.cochlearis (II нейрон)
| |

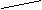

| Nucl.proprii corporis trapezoidei
| |

| Nucl.colliculi inferioris
(IV нейрон)
| |



| Corpus geniculatum mediale
(IV нейрон)
| |
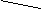

Рис. 21. Пути слухового анализатора
Аксоны клеток дорзального ядра улиткового нерва (II нейронов) направляются по поверхности ромбовидной ямки из латерального угла последней к срединной борозде, образуя мозговые полоски, striae medulleres, отграничивающие с дорзальной стороны мост и продолговатый мозг. В области срединной борозды эти волокна погружаются вглубь, переходят на противоположную сторону и, объединяясь с волокнами трапециевидного тела, образуют латеральную петлю. На пути они вступают в контакт с клетками ядер латеральной петли (III нейроны). Часть аксонов клеток дорзального ядра улиткового нерва, а также ядер трапециевидного тела идут в составе латеральной петли своей стороны.
Таким образом, латеральная петля представляет совокупность аксонов III нейронов пути слухового анализатора, тела которых расположены в верхних оливах, собственных ядрах трапециевидного тела и ядрах латеральной петли. На пути волокна латеральной петли отдают коллатерали к ядрам сетчатой формации. Кроме того, их коллатерали присоединяются к медиальному продольному пучку, посредством которого они осуществляют связь с двигательными ядрами черепных нервов и спинного мозга. Этими связями объясняются рефлекторные ответы (сочетанный поворот головы, глаз и туловища) на слуховые раздражения. Заканчивается латеральная петля в первичных подкорковых слуховых центрах—медиальном коленчатом теле и нижнем бугорке четверохолмия.
Аксоны клеток медиального коленчатого тела (IV нейронов) под названием центрального слухового пути следуют через задний отдел заднего бедра внутренней капсулы в кору средней части верхней височной извилины (извилины Гешля), где заложено ядро слухового анализатора, осуществляющее точный анализ и синтез воспринятых звуковых раздражений.
Переработав полученную информацию, корковый конец слухового анализатора передает ее исполнительной системе, представленной пирамидным и экстрапирамидными путями, осуществляя через них ответную реакцию на звуковые раздражения.
Часть волокон латеральной петли, следующая к нижнему бугорку четверохолмия, переключается на клетки его ядра, аксоны которых, перейдя на противоположную сторону, формируют тектобульбарный и тектоспинальный тракты, заканчивающиеся, соответственно, в двигательных ядрах черепных нервов и передних рогов спинного мозга. По этим путям осуществляются независимые от нашего сознания ответные реакции на неожиданные звуковые раздражения (сторожевой, защитный рефлекс).
В связи с тем, что в каждой латеральной петле идут аксоны клеток слуховых ядер своей и противоположной стороны, одностороннее поражение ее, а также подкорковых центров и коркового ядра слухового анализатора (извилины Гешля) не сопровождается существенным расстройством слуха. Поражение же улиткового нерва и его ядер ведет к потере слуха на своей стороне.








