| Типы реликтов
| Названия растений
|
| Мезофильные реликты полтавской флоры
| Самшит вечнозеленый (Buxus sempervirens) Падуб остролистный (Ilex aquifolium) Папоротники: - птерис критский (Pteris critica) - чистоус величавый (Osmunda regalis) Рододендрон понтийский (Rhododendron ponticum)
|
| Мезофильные реликты тургайской флоры
| Пихта Нордманна (Abies Nordmanniana) Ель восточная (Picea orientalis) Лапина (Pterocarya fraxinifolia) Дуб понтийский (Quercus pontica) Граб обыкновенный (Carpinus betulus) Жимолость-каприфоль (Lonicera caprifolium) Дзелква (Zelkova carpinifolia) Бук восточный (Fagus orientalis) Хмелеграб граболистный (Ostrya carpinifolia) Каштан посевной (Castanea sativa) Липа кавказская (Tilia caucasica)
|
| Гемиксерофильные Средиземноморские реликты
| Лавр благородный (Lanrus nobilis) Земляничник мелкоплодный (Arbutus andrachne) Ладанник крымский (Cistus tauricus) Жимолость тосканская (Lonireca etrusca) Эрика древовидная (Erica arborea)
|
Эта зона богата ценными культурными растениями (маслины, миндаль, инжир, лимоны, мандарины, апельсины, виноград и др.). Она (средиземноморский центр по Н.И. Вавилову) является родиной многих культурных растений (оливковое дерево, рожковое дерево, инжир, миндаль, клевер, капуста и др.).
ЗОНА ПУСТЫНЬ И СТЕПЕЙ УМЕРЕННОГО ПОЯСА
Площадь аридных (от лат. сухой) территорий составляет примерно 48 млн. км2 (или более 30% поверхности суши), где влага является решающим фактором, определяющим биологцческую продуктивность. На рис. 79 представлена мировая карта аридных территорий земного шара, к которым относятся ландшафты пустынь, полупустынь, сухих степей, ксерофитных саванн и др. Выделено 4 биоклиматические зоны: экстраариадная (осадков менее 100 мм), ариадная (годовая сумма осадков 100 - 200 мм), полуаридная (200 - 400 мм) и зона недостаточного увлажнения - субгумидная (более 400 мм). По данным П. Мейгса, на долю экстрааридных территорий приходится 4% суши, аридных - 15%, полуаридных - 14,6% и т.д. Аридные территории распространены на всех материках, причем отношение площади аридных и полуаридных земель к площади континентов составляет: в Австралии - 83%, Африке - 59%, Азии - 38%, Северной и Южной Америке - 10% и 8% соответстенно, а в Европе - всего 1%.
По уточненным данным ЮНЕСКО и ФАО, к категории типичных пустынь относится 23% земной поверхности. На территории бывшего СССР пустыни занимали около 14% его территории (Бабаев и др., 1986). Существует множество классификаций пустынь: по литологии субстрата (каменистые, щебнистые, песчаные, глинистые), по геоботаническим признакам, по характеру растительности, по типам климата (тропические, субтропические, умеренные). В пустынях преобладают сероземные, засоленные типы почв с малым содержанием гумуса.
Во всех пустынях земного шара преобладающими видами растений являются представители таких семейств, как маревые, крестоцветные, сложноцветные, бобовые, парнолистниковые, гречишные и пр. Так, в величайшей пустыне земного шара – Сахаре
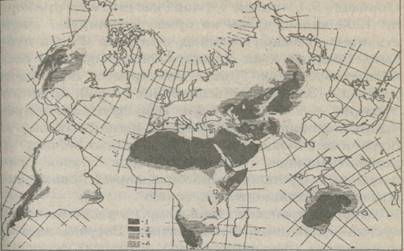
Рис. 79. Аридные территории земного шара (Курьер ЮНЕСКО, №3, 1978).
1 - экстрааридные; 2 - аридные; 3 - семиаридные; 4 - субгумидные.
(площадь 9,1 млн. км2) флора насчитывает примерно 1200 видов, среди которых доминируют представители 6 вышеуказанных семейств. Флора пустынь Средней Азии сравнительно небогата: в песчаных пустынях Турана фиксируется 536 видов растений, Западных Кызылкумов - 533, Южного Устюрта - 256 видов и т.д. Ведущую роль в пустынных ландшафтах играют полынно-маревые и полынные сообщества, а на засоленных почвах галофитные виды (сарсазан, солерос, кокпек, боялыч и др.). Пустыни земного шара имеют свою длительную эволюционную историю.
Вопрос о происхождении флоры и растительности пустынь давно интересовал ученых. Впервые идею о вторичности травянистого типа цветковых растений высказал русский ботаник и географ А.Н. Краснов (1899), считая «эти типы растительности сравнительно новейшими, которые получили развитие в середине третичного периода». Однако, по новейшим палеоботаническим данным отдельные представители травянистых растений появились еще в меловом периоде. Вероятно, во всех случаях возникновение травянистых фитоценозов было связано со многими природными факторами (в основном климатическими), наиболее благоприятными для возникновения и формирования открытых ландшафтов.
По вопросу происхождения растительности пустынной зоны имеется ряд гипотез. Так, весьма интересна гипотеза обширных миграций М.Г. Попова (1927), по которой формирование пустынной флоры началось еще в меловом периоде. Центр возникновения и развития первичной пустынной растительности земного шара располагался в южном полушарии - на материке ГонДвана. Современная флора пустынь образовалась из двух исходных флор: мезофильной лесной флоры Гинкго и ксерофильной флоры Вельвичих, которые разделялись древним океаном - Тетис. Первая - существовала на северном побережье, вторая - на южном побережье Тетиса. В миоцене после спада вод Тетиса и выхода на земную поверхность из-под моря обширных участков суши, именно эти две флоры начали взаимодействовать между собой, в результате чего сформировалась ксерофильно-пустынная флора. М.Г. Попов основывался на палеоботанических работах А. Энглера, который относил начало ксерофильной флоры к меловому периоду. Именно к флоре Вельвичих принадлежали различные виды маревых, пар нолистниковых, гребенщиковых, эфедры и др., которые в настоящее время встречаются во флоре пустынь всех континентов.
Однако большинство отечественных исследователей (В.Л. Комаров, И.М. Крашенинников, М.М. Ильин и др.) главное значение придавали автохтонному развитию флоры и растительности пустынь умеренного пояса. Следует отметить, что впервые В.А. Комаров отметил роль древних литоралей в формировании пустынных фитоценозов. На основе изучения флоры Монголии, он сделал вывод, что формирование пустынной флоры Центральной Азии происходило в основном за счет усыхания древних водоемов, на литоралях которых и формировались пустынные ценозы (с участием галофитных видов маревых).
М.М. Ильин (1958, с. 144) писал: «Аридные условия в Центральной Азии существовали по крайней мере с верхнего мела, не претерпевая никаких острых моментов, которые могли бы нарушить непрерывное развитие ксерофильных флор этих обширных территорий: пустыни Средней и Центральной Азии разновозрастны, причем каждая из этих территорий имеет свою собственную историю». Он наметил следующие этапы ксерофитизации растительности пустынь Евразии. Так, к позднему мелу относится зарождение пустынного ландшафта Центральной Азии. В более северных районах, в частности в Средней Азии, опустынивание началось значительно позже - с палеогена. Здесь при нарастающей аридизации климата создавались благоприятные условия для формирования пустынных формаций, причем это происходило главным образом за счет литоралей с засоленными почвогрунтами, образовавшимися после спада вод древнего океана - Тетис. Затем процесс ксерофитизации растительности из Средней Азии стал распространяться на север, захватив в миоцене территорию Казахстана и юга Русской равнины. Позднее, в плиоцене и плейстоцене, вследствие похолодания климата и тектонических процессов, происходила дальнейшая эволюция пустынной флоры, которая приобретала все более современный облик. Для сравнения укажем, что мексиканские и отчасти американские пустыни (Мохаве, Колорадо, Долина Смерти и др.) имеют также миоценовый возраст. Однако находки эоценовой опунции свидетельствуют о еще более древних корнях пустынной флоры пустынь Северной Америки.
В результате периодической смены климатов Земли менялись и границы пустынных территорий. Например, пески Сахары в определенные периоды (около 20 тыс. лет назад)'простирались на 5 - 6° (примерно 400 км) южнее их современных границ. По имеющимся материалам, полуаридные и полугумид-ные территории тропической Африки, Аравийского полуострова и Иранского нагорья примерно 12 - 7 тыс. лет назад были более влажными, чем в настоящее время. Однако в нынешнем столетии на аридные территории стал оказывать существенное влияние антропогенный фактор. По оценке ЮНЕП, в мире ежегодные потери только орошаемых земель в результате опустынивания составляют б млн. га. К настоящему времени площадь антропогенных пустынь составила 9 млн. км2 (примерно равна площади Сахары). Поэтому данные об эволюции флоры и растительности пустынной зоны весьма необходимы для успешного решения проблемы охраны аридных территорий земного шара.
Происхождение флоры и растительности степной зоны. Параллельно с формированием пустынь происходило образование открытых степных ландшафтов. По этому поводу акад. И.П. Герасимов (1976) писал: «Основным направлением в эволюции природы зональных географических ландшафтов в течение кайнозоя, было постепенное обособление из лесных ландшафтов, господствовавших в начале кайнозоя, безлесных, травянистых ландшафтов».
В географической литературе вопрос о причинах безлесия степей обсуждался давно. Допускались различные причины: климатические, геоморфологические, почвенные, антропогенные и др., но одним фактором (климат, почвы, равнинность рельефа и пр.) нельзя было решить данную проблему. Значительный вклад в решение так называемого «степного вопроса» внесли результаты споро-пыльцевых исследований неогеновых отложений юга Русской равнины.
По данным Е.Н. Анановой (1966), распространение открытых степных ландшафтов на юге Русской равнины началось с позднего миоцена (верхний сармат), что было обусловлено комплексом физико-географических факторов. Также большое влияние на формирование степной зоны оказало поднятие и оформление Кавказа как горной системы, происшедшее в конце среднего - верхнем сармате. Это способствовало усилению континентальноети и аридности климата на юге Русской равнины. В таких условиях наиболее фитоценотически сильным оказался травянистый тип растительности.
Результаты палинологических исследований верхнеплиоценовых и четвертичных отложений Прикаспийской низменности показали, что с позднего плиоцена здесь начали господствовать открытые полупустынностепные ландшафты со значительным участием ксерофитов и галофитов. Среди пыльцы маревых было обнаружено 44 вида, обитающих на мокрых и пухлых солончаках, на солонцах и солончаках (Вронский, 1970). Все это свидетельствует о том, что во время регрессий Каспийского моря, когда от воды освобождались обширные участки суши с засоленными почвогрунтами, на них интенсивно происходило формирование как полупустынных, так и степных типов растительности.
Таким образом, в кайнозое (палеоген, неоген) на равнинах юга Евразии возникли и широко распространились пустынные и степные формации, в которых доминирующими жизненными формами растений стали однолетние и многолетние травы из семейств: маревых, сложноцветных, злаковых, бобовых, крестоцветных, лилейных, зонтичных, гвоздичных и др. В результате длительной хозяйственной деятельности человека отмечены коренные изменения степных ландшафтов не только Евразии, но и других континентов (аргентинская пампа, североамериканские прерии). Так, за последние 100 лет в типичных черноземах Воронежской области в пахотном слое содержание гумуса уменьшилось с 10 – 13% до 7%. В некоторых районах степной зоны России отмечена значительная деградация пастбищ из-за чрезмерного выпаса скота и т.д. Поэтому в последние годы за рубежом и в нашей стране развернуты широкие исследования по экологической оптимизации степного природопользования с целью снижения антропогенных нагрузок на агроэкосистемы степной зоны.
ЗОНА ТАЙГИ
Зона бореальных хвойных лесов умеренного пояса северного полушария занимает обширные пространства Евразии и Северной Америки (см. рис. 77). Ее площадь составляет 13,4 млн. км2, из них 9 – в России. Тайга является в нашей стране наиболее крупной по площади природной зоной, при этом горная тайга занимает более 50% всей площади России.
По сравнению с тундровой зоной тайга получает значительно больше тепла. Лето там прохладное, но достаточно теплое для произрастания хвойных лесов. Резко увеличена длительность безморозного периода: 75 – 90 дней на севере зоны и 100 – 120 дней на юге. Годовая сумма атмосферных осадков колеблется в пределах 300 – 800 мм, причем значительная их часть выпадает в виде снега. В тайге распространена многолетняя мерзлота (особенно к востоку от Енисея), которая играет решающую роль в распределении типов почв и растительности. Многолетняя мерзлота имеет широкое распространение и в Северной Америке, причем в Канаде она занимает более половины бореальной зоны, способствуя сильному заболачиванию территории. Господствующим типом почв здесь являются подзолистые. Биомасса растений таежной зоны составляет 1500-3000 ц/га.
Доминирующая жизненная форма растений в тайге представлена фанерофитами – вечнозелеными хвойными деревьями (ель, сосна, пихта, кедр, лиственница).
Наиболее типичным проявлением таежного типа растительности является темнохвойная тайга – густой, тенистый, вечнозеленый лес, образованный в основном елями и пихтами с участием теневыносливых видов сосен. Наибольшую площадь в России занимают лиственничные формации (лиственница сибирская, Сукачева, даурская) 269 млн. га (38% всей лесопокрытой площади). В Северной Америке таежные формации (в Канаде и США) представлены темно-хвойными лесами из различных видов елей (черной, Энгельмана, канадской) с примесью пихты бальзамической и лиственницы американской. На Аляске (плато Юкон) леса таежного типа состоят из лиственниц, елей (черной, ситхинской) и тсуги западной.
Вопрос о происхождении флоры и растительности таежной зоны имеет свою историю и до настоящего времени окончательно не решен. Имеется несколько точек зрения на проблему генезиса растительности тайги. Интересная гипотеза была высказана акад. В.Б. Сочавой (1944), который полагал, что развитие растительности таежной зоны происходило автохтонно в пределах современной зоны путем постепенного выпадения из состава арктотретичных лесов более теплолюбивых древесных пород. Следует отметить, что флоры из миоценовых отложений Арктики получили название – арктотретичные (по А. Энглеру). Более разнообразные по составу арктотретичные леса постепенно преобразовывались в простые монодоминантные фитоценозы. Как равнинная, так и горная тайга образования сравнительно молодые.
М.Г. Попов (1949) на основе изучения ископаемых флор предложил гипотезу приполярного происхождения тайги:
1. тайга – молодое образование, связанное с деградацией арктотретичных лесов;
2. область первоначального формирования тайги лежала к северу от области современного ее распространения, в пределах ныне безлесных пространств Крайнего севера;
3. распространение тайги в пределах умеренного пояса протекало в основном с севера на юг;
4. тайга горных районов, расположенных к югу от таежной зоны, - явление вторичное, более позднее.
А.И. Толмачев (1954) предложил гипотезу горного происхождения тайги, по которой она первоначально возникла в горных условиях, а затем по мере деградации арктотретичных лесов начала занимать равнинные пространства севера Евразии. При этом скорость миграции хвойных пород из горных районов на равнины составляла 100 – 150 м/год. Однако новейшие исследования биоэкологических свойств древесных пород по скорости расселения и миграции показали, что эти процессы проходят значительно медленнее. По расчетам И.Ф. Удры (1988), скорость миграции темно-хвойных эдификаторов тайги (ель, пихта, кедр) составляет всего 10 – 25 м/год, а для светлохвоиных (сосна обыкновенная, лиственница) – до 30 – 60 м/год.
Отметив лишь основные концепции происхождения флоры и растительности таежной зоны, видно, что эта проблема довольно сложна и для ее окончательного решения необходимы палеоботанические исследования неогеновых отложений севера Евразии и Северной Америки. В последние годы получены интересные материалы по спорово-пыльцевым исследованиям неогеновых отложений Русской равнины (Нижняя Кама, Поволжье, Белоруссия и др.). Это позволило внести значительный вклад в решение вопроса о происхождении таежной растительности.
По данным Е.Н. Анановой (1966) возникновение тайги и ее эволюция протекала следующим образом:
1. формирование таежных лесов как нового зонального типа растительности происходило в результате длительного процесса эволюции флоры и растительности одновременно на больших пространствах;
2. наиболее, вероятно, что темнохвойные формации тайги образовались автохтонно, но обеднение их миоценовыми реликтами происходило в северных районах зоны раньше, чем в южных;
3. становление таежного типа растительности явление относительно молодое, связанное с неогеном;
4. горная тайга - вторичное образование.
Палеоботанические материалы позволили сделать вывод о том, что возникновение таежной зоны на севере и степной на юге Русской равнины взаимосвязаны.
Однако в последние десятилетия на естественный ход эволюции таежной растительности Евразии и Северной Америки сильное воздействие стали оказывать антропогенные факторы. Так, рубка леса, гидротехническое и промышленное строительство привели к резкому уменьшению лесных ресурсов зоны. Страдают таежные ландшафты в России и Канаде от вредных выбросов (соединения серы, никеля, меди, цинка, марганца, кобальта и др.) промышленных центров, в частности на Кольском полуострове вокруг них возникли участки полностью деградированной тайги. Помимо интенсивных лесозаготовок в тайге расширились геологоразведочные работы (нефть, газ, уголь). В качестве негативных последствий всего комплекса антропогенных факторов отмечается сокращение запасов деловой древесины, ухудшение лесорастительных условий, уменьшение биологической продуктивности лесов, снижение урожайности грибов, ягод и количества промысловых ценных животных. Поэтому в России и Канаде начаты лесовосстановительные работы в таежной зоне, а также мероприятия по увеличению площади охраняемых природных территорий. Все это позволит сохранить таежные ландшафты – как основной источник пополнения запасов кислорода в атмосфере нашей планеты.
ЗОНА ТУНДРЫ
Занимает северные окраины Европы и Северной Америки, причем ее южная граница примерно совпадает с июльской изотермой +10°С. Для тундровой зоны характерны низкие температуры воздуха, короткий вегетационный период, присутствие многолетней мерзлоты, незначительное количество атмосферных осадков (200 - 400 мм), преобладание глеево-болотных почв. Биомасса растений тундры составляет 40 – 280 ц/та при годовом приросте в 10 – 25 ц/га.
Флора тундры насчитывает всего 250 500 видов; растительный покров образован главным образом мхами, лишайниками и травянистыми растениями с участием низкорослых кустарников. Значительно распространены, стелющиеся и подушкообразные жизненные формы растений. Немногочисленные травы (осоки, полярные маки, одуванчики) и кустарники (голубика, морошка, брусника) чередуются с карликовой березкой, полярной ивой, кедровым тлаником. В Северной Америке кустарничковые тундры определяют ландшафт больших пространств и образованы в основном растениями из семейства вересковых.
В настоящее время большинство ученых считают безлесие тундры зональным явлением, обусловленным комплексом неблагоприятных для леса факторов, среди которых ведущая роль принадлежит низким температурам воздуха в сочетании с высокой относительной влажностью. Ведь еще М.В. Ломоносов отмечал, что «тундрами называются места, заросшие мхом».
Постепенно подготавливались условия и для образования растительности тундровой зоны. Ископаемые остатки тундровой флоры обнаруживаются лишь в четвертичных отложениях. Эта безлесная ландшафтная зона Евразии начала свое формирование именно в плейстоцене. По данным А.И. Толмачева (1986), образование тундровой флоры происходило первоначально не на равнинах севера Евразии, а в горах Северо-Восточной Сибири, где общее похолодание климата прежде всего сделало невозможным дальнейшее произрастание лесных формаций. Затем формирование тундровых ландшафтов протекало на равнинных пространствах Восточной Сибири, чему способствовало наличие здесь в четвертичном периоде территорий, не покрытых ледниками. Постепенно тундровая флора стала распространяться зонально, образовав в конце концов самую новейшую ландшафтно-ботаническую зону Евразии. Особенно тундра молода в секторах Арктики, которые в плейстоцене были покрыты льдом, поэтому здесь тундра имеет даже послеледниковый возраст. Характерным элементом тундровой зоны, как и таежной, являются моховые бореальные болота, которые начали широко распространяться только с четвертичного периода.
Дальнейшая эволюция флоры тундровой зоны оказала существенное влияние на ее современный облик растительности. Так, палеоботанические исследования послеледниковых торфяников тундровой зоны (Ямал, низовья р. Лены, Чукотка) зафиксировали находки пыльцы карликовой березки (Betula папа), ольхи кустарниковой (Alnus fruticosa), сосны обыкновенной (Pinus silvestris), ели, лиственницы, тополя и др. Обнаружены остатки некоторых вечнозеленых растений: жимолости (Lonicera edulis), рододендрона (Rhododendron chrysantum), можжевельника сибирского (Yuniperus cedrus), багульника (Ledum palustre), брусники (Vaccinium vitis), черники (Vaccinium myrtillus), Кассиопеи (Cassiope tetragona), морошки (Rubus chamaemorus), андромеды (Andromeda polifolia) и др. Многие виды растений, несмотря на дальнейшее похолодание климата, остались в тундре и произрастают там до настоящего времени. Эти реликты лесного прошлого являются молодыми элементами, связанными с термическим оптимумом голоцена (послеледниковья). Как считает Б.А. Тихомиров, именно этим палеогеографическим фактором и можно объяснить возможность произрастания в тундровой зоне вечнозеленых кустарников и кустарничков (брусника, пушица, андромеда, багульник и др.), свойственных болотным ассоциациям лесной зоны.
Однако в последние десятилетия сильно возросло антропогенное воздействие на природные ландшафты тундровой зоны. Ускоренными темпами идет антропогенная трансформация типичных тундр полуострова Ямал под воздействием ряда газоконденсатных месторождений. Уменьшается общее покрытие растительного покрова и количество видов растений, упрощается структура фитоценозов и т.д. Чрезмерный выпас северных оленей в некоторых районах тундровой зоны привел к полному разрушению лишайникового покрова и к деградации пастбищ. Растительность тундровой зоны с учетом природных особенностей (замедленный биологический круговорот, низкая продуктивность и пр.) отличается повышенной чувствительностью к различным атмосферным загрязнителям (Вронский, 1996). Кроме того, экосистемы тундры обладают весьма малой способностью к самоочищению. Все это необходимо учитывать при освоении и разумном использовании природных ресурсов тундровой зоны.
Таким образом, в кайнозое под влиянием комплекса природных факторов (в основном климатических) возникали один за другим новые типы растительности, образовавшие горизонтальные ботанико-географические зоны суши. Однако, значительная часть природных зон характеризуется вертикальной дифференциацией ландшафтов, которая в горных странах перерастает в высотную поясность.
ПРОИСХОЖДЕНИЕ ВЕРТИКАЛЬНОЙ РАСТИТЕЛЬНОЙ ПОЯСНОСТИ
Высотная поясность ландшафтов – закономерная смена природных экосистем в горных районах, обусловленная высотой над уровнем моря. Она связана с понижением температуры воздуха, увеличением количества ультрафиолетовой радиации, снижением величины испаряемости и т.д. Вертикальная поясность наиболее ярко выражена в смене климата, почв и типов растительности по высоте. Высотная поясность фитоценозов во многих отношениях сходна с широтным распределением основных типов растительности континентов (рис. 80).

Рис. 80. Соотношение между высотным и широтным распределением основных типов растительности суши (по Ф. Рамаду, 1981).
Эта растительность по ряду признаков имеет сходство с тундровой: общие жизненные формы (стелющиеся и подушкообразные), значительная продолжительность жизни растений, преобладание подземной части в биомассе (63 – 90%) и др. Так, в высокогорьях стелющуюся форму роста имеют различные крупки, астрагалы, змееголовники; они плотно прижаты к прогреваемому солнечными лучами щебню и тем самым спасаются от замерзания и сильного ветра. В горах встречаются такие типы растительности, как нагорные ксерофиты, холодные сухие пустыни, пуна и др., отсутствующие на равнинах.
Однако флора горных районов всегда богаче флоры прилегающих равнинных территорий. Это связано с сохранением в горных убежищах реликтов, с высотной поясностью и обилием горных изолятов, в которых интенсивно протекают процессы видообразования растений. Для высокогорных флор характерна значительная роль эндемичных видов растений. Так, флора Кавказа насчитывает 6350 видов, из которых 25% эндемичны. Флора горных районов Средней Азии оценивается в 5500 видов, из них 25 – 30% - эндемичные формы, в то время как на равнинах южных пустынь насчитывается около 2000 видов растений (Агаханянц, 1986).
В конце XVIII в. А. Гумбольдт в горах Южной Америки установил наличие вертикальных поясов растительного покрова. Крупнейшие ботаники (Геер, Энглер, Краснов, Вульф и др.) придавали исключительное значение изучению высокогорных флор, так как это служило ключом для раскрытия общих закономерностей истории формирования растительности земного шара. Явление вертикальной поясности особенно полно представлено в горах экваториально-тропической зоны, где наблюдается полный набор растительных поясов – от влажнотропических лесов до альпийских лугов. Знаменитый географ А.Н. Краснов (1882 – 1914), описывая высокогорную флору о. Ява, отмечал наличие ряда растений (восковник, голубика, брусника, морошка, рододендрон и др.), которые в настоящее время обитают на бореальных болотах севера Евразии. Он писал: «Эта близость флоры горных вершин Явы к столь противоположным по свойствам тундрам севера представляет огромный интерес, так как она показывает, что флора тундры может быть непосредственным производным флоры тропического леса». Иначе говоря, на горных вершинах о. Ява наблюдается своеобразный, но еще не зашедший далеко процесс перерождения тропических лесов в альпийскую растительность. Явление вертикальной ботанико-географической поясности в целом значительно древнее. Однако современный горный рельеф материков создан в основном движениями альпийской фазы орогенеза, что затрудняет изучение вертикальной поясности более древних геологических периодов.
По данным Е.В. Вульфа (1944), в Средиземноморье вертикальная поясность растительности существовала во всяком случае с конца эоцена. Так, по палеоботаническим данным в Родопских горах (Болгария) в эоцене выделялись следующие вертикальные пояса растительности (снизу вверх): пояс из дуба, земляничного дерева, маслины, кипариса и др.,
- пояс из дуба, клена, ясеня и граба,
- пояс из ели, пихты, сосны, ивы, бука, березы,
- пояс из рододендрона, брусники, голубики и др. (альпийский).
По материалам В.П. Малеева (1941), на Кавказе в неогене располагались такие вертикальные растительные пояса: в самом низу – средиземноморцы и реликты полтавской флоры, выше располагался пояс дубового и букового леса, - пояс темнохвойного леса, - субальпийский и альпийский пояса.
Большинство исследователей признают автохтонное развитие высокогорной флоры Кавказа, на которое значительное влияние оказали тектонический фактор и четвертичные оледенения, приведшие к обогащению флоры бореальными и арктическими элементами. Вышеуказанная вертикальная поясность растительности Кавказа в неогене имеет значительные черты сходства с современными высотными поясами, особенно Западного Закавказья. Здесь отмечаются (снизу вверх): влажные субтропические леса, пояс дубовых, затем буковых и смешанных буково-хвойных лесов, которые сменяются субальпийскими (высокотравными лугами и зарослями из стелющихся рододендронов) и альпийскими (низкотравными) лугами.
Высокогорная растительность формировалась по мере роста горных сооружений, путем видоизменения первоначальной растительности данной территории с учетом местных природных условий. А.И. Толмачев (1986), на территории Евразии выделяет 4 основных типа высокогорной растительности:
1. Альпийский ландшафт (Альпы, Кавказ, Тянь-Шань, Алтай и др.). Характерными растениями являются мезофиты (злаковые, осоки, разнотравье). Эта флора возникла из горной мезофильной растительности различного типа (осветленные леса, луга), существовавшей первоначально в лесном окружении.
2. Гольцовый (горнотундровый) ландшафт (горы Восточной Сибири), приспособленный к суровым малоснежным зимам. Заросли кустарников и кустарничков (ивы, кедровый стланец, рододендроны, дриада, вересковые и пр.). Генетически этот тип растительности связан с мезофильными лесами северо-востока Азии.
3. Ландшафт нагорных ксерофитов (горы юга Средней Азии, Афганистана, Ирана, Армянского нагорья, Балканского полуострова и др.). Преобладают жестколистные, колючие, подушкообразные кустарники и полукустарники, многолетние травы (астрагал, сложноцветные, зонтичные, губоцветные и др.). Генетически нагорные ксерофиты тяготеют к ксе-рофитным типам растительности древнего Средиземноморья, связанных с группировками частично облесенных горных ландшафтов.
4. Нагорные степи и высокогорные пустыни Азии. Характерны ксерофитные многолетние травы (дерновидные злаки, полыни, терескен и др.). Генетически эти формации возникли из степной растительности внутренних областей Азии.
Как считает В.Б. Сочава, альпийская флора возникла автохтонно, т.е. «на месте», причем ее ядро сложилось в сравнительно древних, еще мезозойских горах востока Азии и запада Северной Америки. Причем необходимо разграничивать альпийскую и арктическую (тундровую) флоры, каждая из которых имеет свое автохтонное ядро, но более древнее у альпийской флоры. Эти флоры взаимодействовали путем взаимной миграции различных видов растений, поэтому в Альпах насчитывается до 30% видов, общих с тундровой флорой.
В последние годы возросли антропогенные нагрузки (сведение лесов, перепас скота, загрязнение атмосферы и др.) на высокогорные экосистемы. Так, уничтожение реликтовых лесных формаций в горах Средней Азии послужило причиной значительного обезлесивания горных ландшафтов, что в свою очередь привело к аридизации климата и ксерофитизации растительности. Аналогичные процессы происходят и на Кавказе, где наблюдается деградация высокогорной растительности, особенно вблизи крупных промышленных центров и рекреационных зон.
Таким образом, растительный покров суши прошел длительную эволюцию своего развития, в результате которой сформировались современные типы растительности Земли. Однако появление человека и возрастающее его воздействие на биосферу, стало оказывать существенное влияние на естественный ход эволюции растительного покрова (селекция, создание новых культурных видов растений и др.). Но в последние десятилетия антропогенный фактор стал главной причиной катастрофического исчезновения лесных формаций с лика Земли. В отчете ЮНЕП «О состоянии окружающей среды к 2000 году» отмечено, что «сведение лесов – вероятно, наиболее серьезная экологическая проблема, стоящая перед человечеством». В настоящее время ежегодно сводится до 16,8 млн. га влажнотропических лесов.
Значительное воздействие на площади природных зон (тундра, тайга, степи и др.) в будущем может оказать глобальное потепление климата. Для оценок предстоящих изменений ландшафтов создаются модели с привлечением палеогеографических материалов. Так, сопоставление карт современной растительности и палеорастительности для оптимума голоцена (потепление климата на 1,4°С) и для микулинского межледниковья (повышение температуры на 2,2°С) показало наличие как совпадений, так и некоторых расхождений в распределении ландшафтных зон. Например, при глобальном потеплении климата на 1,4°С (ожидается на 2000 г.) на территории бывшего СССР площади тундры и хвойных лесов сократятся, примерно, в 2 – 3 раза, но зато в 5 раз возрастет ареал широколиственных лесов и т.д. (Кобак, Кондрашева, 1992). Однако представленные модели имеют довольно условный характер, так как в них (и в зарубежных) не учитывается важный фактор изменения режима увлажнения. Такие исследования важны для оценок развития природных ландшафтов и для прогноза формирования агроэкосистем в условиях парникового эффекта.
Перед человечеством возникла сложная проблема экологического плана – не только сохранить, но и увеличить площади лесов на земном шаре, которые создают благоприятную природную среду для здоровой жизни будущих поколений. Как напоминание звучат слова из предисловия ж. «Курьер ЮНЕСКО» (1989) «Хрупкий лес»: «В конечном счете мы живем за счет растений, особенно деревьев, - основных производителей кислорода, необходимого для существования рода человеческого. И если мы не сможем сохранить древесную растительность, то рано или поздно погибнем вместе с ней».
Глава XI. ПРОИСХОЖДЕНИЕ ЧЕЛОВЕКА И ЕГО ВЛИЯНИЕ НА БИОСФЕРУ
Человек - существо свободное, поелику одарено разумом и свободною волею: Что свобода его состоит в избрании лучшего, что сие лучшее познает он и избирает посредством разума, постигает пособием ума и стремится всегда к прекрасному, величественному, высокому...
А. И. Радищев
ДРЕВНОСТЬ ЧЕЛОВЕКА
В природных условиях в палеонтологической летописи останки человека сохраняются очень плоха. Значительно лучше сохраняются орудия, которыми пользовался первобытный человек, поскольку часто они изготовлялись из твердого - каменного материала. Таким образом обнаруженные в слоях почвы или в пещерах каменные орудия, естественно, могли дать первую косвенную информацию о существовании людей в прошлые эпохи. Грубые каменные орудия труда и охоты, изготовленные нашими далекими предками, представляют собой исторические свидетельства, по которым мы узнаем о культурной жизни доисторического человека. В течение довольно долгих лет, эти орудия, изготовленные из кремня, сланца, обсидиана и других пород, считались всего лишь причудами природы. Только на исходе XVI века Михаэль Мерканти, личный врач папы Климента VIII, угадал истинную природу этих предметов. Во многих случаях изделия древнего человека встречались в пещерах, наряду с останками ископаемых животных: первобытного слона, носорога, гиены и медведя. В Англии преподаватель геологии Оксфордского университета священник Вильям Бекленд (1784 - 1853) в Павиландской известковой пещере на побережье Уэлса обнаружил целые залежи доисторических предметов: кремниевые орудия, разные украшения и инструменты из кости и бивня слона лежали вперемежку с останками животных тех же видов, которые были ранее найдены в других пещерах Западной Европы. Там же был обнаружен человеческий скелет.
Несколько позже католический священник Джон Макэнери обследовал огромную извилистую известковую пещеру (пещера Кента), находящуюся на девонском побережье Англии. Он нашел каменные орудия антропогенного происхождения возле останков тех же вымерших животных, обнаруженных В. Беклендом в Павилендской пещере. Несмотря на суровые осуждения церкви и даже самого В. Бекленда, Макэнери продолжал считать сделанные им открытия неоспоримым свидетельством того, что человек жил в Англии задолго до библейского потопа и был современником вымерших животных, останки которых он обнаружил.
Наконец, выдающийся английский геолог Ч. Лай-ель в своей книге «Древность человека» (1864) привел веские доказательства в пользу того, что орудия труда древнего человека могут указывать на степень его древности. Возраст этих грубо обработанных камней определялся по условиям их залеганий на разных глубинах совместно с ископаемыми животными.
Ч. Дарвин рассматривал человека как результат эволюции животного мира. Его обширный труд «Происхождение человека и половой отбор» был выпущен в 1871 г. Однако в то время, по существу, еще не были известны иско






