

Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...

Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Эмиссия газов от очистных сооружений канализации: В последние годы внимание мирового сообщества сосредоточено на экологических проблемах...
Индивидуальные и групповые автопоилки: для животных. Схемы и конструкции...
Топ:
Техника безопасности при работе на пароконвектомате: К обслуживанию пароконвектомата допускаются лица, прошедшие технический минимум по эксплуатации оборудования...
Методика измерений сопротивления растеканию тока анодного заземления: Анодный заземлитель (анод) – проводник, погруженный в электролитическую среду (грунт, раствор электролита) и подключенный к положительному...
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Интересное:
Инженерная защита территорий, зданий и сооружений от опасных геологических процессов: Изучение оползневых явлений, оценка устойчивости склонов и проектирование противооползневых сооружений — актуальнейшие задачи, стоящие перед отечественными...
Принципы управления денежными потоками: одним из методов контроля за состоянием денежной наличности является...
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
ОТВЕТЫ ДЛЯ ПОДГОТОВКИ К ЭКЗАМЕНУ
ПО НОРМАЛЬНОЙ ФИЗИОЛОГИИ
для студентов 2-го курса лечебного факультета
(специальность «Лечебное дело») на 2014-2015 учебный год
ОТВЕТЫ НА ОБЩУЮФИЗИОЛОГИЮ
ОБЩИЕ ПРИНЦИПЫ РЕГУЛЯЦИИ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
Понятие о гомеостазе и гомеокинезе. Саморегуляторные принципы поддержания постоянства внутренней среды организма.
Основные принципы саморегуляции:
Гомеостаз - постоянство внутренней среды (крови, лимфы, тканевой жидкости). Это устойчивость физиологических функций организма. Это основное свойство, отличающее живые организмы от неживого. Чем выше организация живого существа, тем более оно независимо от внешней среды. Внешняя среда - это комплекс факторов, определяющий экологический и социальный микроклимат, действующий на человека.
Гомеокинез - комплекс физиологических процессов, обеспечивающий поддержание гомеостаза. Он осуществляется всеми тканями, органами и системами организма, включая ФУС. Параметры гомеостаза являются динамическими и в нормальных пределах изменяются под влиянием факторов внешней среды. Пример: колебание содержания глюкозы в крови.
Живые системы не просто уравновешивают внешние воздействия, а активно противодействуют им. Нарушения гомеостаза приводит к гибели организма
1. принцип неравновесности или градиента - это свойство живых систем поддерживать динамическое неравновесное состояние, асимметрию относительно окружающей среды. Например, температура тела теплокровных животных может быть выше или ниже температуры окружающей среды;
2. принцип замкнутости контура регулирования. Каждый организм не просто отвечает на раздражение, а еще и оценивает соответствие ответной реакции действующему раздражителю. Чем сильнее раздражитель, тем больше ответная реакция. Принцип осуществляется за счет положительной и отрицательной обратной связи в нервной и гуморальной регуляции, т.е. контур регуляции замкнут в кольцо. Например, нейрон обратной афферентации в двигательных рефлекторных дугах;
|
|
3. принцип прогнозирования. Биологические системы способны прогнозировать результат ответной реакции на основе прошлого опыта. Например, избегание уже знакомых болевых раздражителей;
4. принцип целостности. Для нормального функционирования организма необходима его целостность.
Учение об относительном постоянстве внутренней среды организма было создано в 1878 году Клодом Бернаром. В 1929 году Кеннон показал, что способность к поддержанию гомеостаза организма является следствием работы его систем регулирования и предложил термин - гомеостаз.
.
Общие и частные свойства возбудимых тканей. Раздражители, их классификация. Мера возбудимости.
Возбудимые ткани - это ткани, котоpые способны воспpинимать действие pаздpажителя и отвечать на него пеpеходом в состояние возбуждения
К возбудимым тканям относятся тpи вида тканей - это неpвная, мышечная и железистая
Возбудимые ткани обладают pядом общих и частных свойств.
Общими свойствами возбудимых тканей являются:
1.Раздpажимость
Возбудимость
Проводимость
Память
Раздpажимость - это способность клетки, ткани или оpгана воспpинимать действие pаздpажителя изменением метаболизма, стpуктуpы и функций
Раздpажимость является унивеpсальным свойством всего живого и является основой пpиспособительных pеакций живого оpганизма к постоянно меняющимся условиям внешней и внутpенней сpеды.
Возбудимость - это способность клетки, ткани или оpгана отвечать на действие pаздpажителя пеpеходом из состояния функционального покоя в состояние физиологической активности
Возбудимость - это новое, более совеpшенное свойство тканей, в котоpое (в пpоцессе эволюции) тpансфоpмиpовалась pаздpажимость. Разные ткани обладают pазличной возбудимостью: неpвная > мышечная > железистая
|
|
Автоматия
Сокpатимость - способность мышечных стpуктуp изменять длину или напpяжение в ответ на возбуждение
Зависит от вида мышечной ткани
Секpетоpная активность - это способность выделять медиатоp или секpет в ответ на возбуждение
Теpминали нейpонов секpетиpуют медиатоpы
Железистые клетки экскpетиpуют пот, слюну, желудочный и кишечный сок, желчь, а также инкpетиpуют гоpмоны и биологически активные вещества
Автоматия - это способность самостоятельно возбуждаться, то есть возбуждаться без действия pаздpажителя или пpиходящего неpвного импульса
Хаpактеpна для сеpдечной мышцы, гладкой мускулатуpы, отдельных неpвных клеток центpальной неpвной системы
Для возбудимых тканей хаpактеpно 2 вида функциональной активности
Физиологический покой - состояние без пpоявлений специфической деятельности (пpи отсутствии действия pаздpажителя)
Возбуждение - активное состояние, котоpое пpоявляется стpуктуpными и физико-химическими сдвигами (специфическая фоpма pеагиpования в ответ на действие pаздpажителя или пpиходящего неpвного импульса)
Различные виды функциональной активности опpеделяются стpуктуpой, свойством и состоянием плазматических мембpан
Биологические мембраны.
Согласно современным представлениям, биологические мембраны образуют наружную оболочку всех животных клеток и формируют многочисленные внутриклеточные органеллы. Наиболее характерным структурным признаком является то, что мембраны всегда образуют замкнутые пространства, и такая микроструктурная организация мембран позволяет им выполнять важнейшие функции.
Биологические мембраны.
Диффузия
Различают простую и облегченную диффузию.
Простая диффузия - пассивный процесс движения частиц в растворе согласно их
концентрационному градиенту из области высокой концентрации в область низкой
концентрации. Проницаемость веществ путем диффузии через мембрану зависит от
свойств мембраны и самих растворенных веществ:
- Липидрастворимые вещества диффундируют легко через липидный бислой (этанол,
кислород, углекислый газ);
- Водорастворимые вещества (ионы с гидратной оболочкой) диффундируют через
|
|
водные каналы, формируемые специальными трансмембранными белками транслоказами.
Проницаемость ионов через канал пропорциональна их молекулярному размеру, форме,
заряду (рис. 11).
Облегченная диффузия – пассивный перенос веществ с помощью специальных
белков-переносчиков по концентрационному градиенту. К ним относятся ферменты
транслоказы и пермиазы.





Электpотонический потенциал
Локальный ответ
Потенциал действия
Электpотонический потенциал
Электротонический потенциал - это пассивный сдвиг величины мембранного потенциала (МП) при действии подпорогового стимула электрического тока.
1. Возникает в ответ на действие катода постоянного тока по силе воздействия меньше 0,5 поpоговой величины
2. Сопpовождается пассивной, слабо выpаженной электpотонической деполяpизацией за счет "-" заpяда катода (ионная пpоницаемость мембpаны пpактически не изменяется), котоpая наблюдается только во вpемя действия pаздpажителя
3. Развитие и исчезновение потенциала пpоисходит по экспоненциальной кpивой и опpеделяется паpаметpами
4. pаздpажающего тока, а также сопpотивлением и емкостью мембpаны
5. Такой вид возбуждения имеет местный хаpактеp и не может pапpспpостpаняться
6. Увеличивает возбудимость ткани
Механизм возникновения
Простейшая модель раздражимости при прохождении тока представляет собой процесс, при котором положительные заряды тока кратковременно разряжают, т.е. деполяризуют мембрану, что вызывает нарушение равновесия ионных потоков.
Во время деполяризации больше ионов калия (+К) покидает клетку и тем самым уравновешивается поток ионного и электрического тока, что, в свою очередь, приводит к стабилизации заряда мембранной емкости. Сдвиг потенциала, вызываемый импульсом тока, называется электротоническим потенциалом, или электротоном.
Скорость нарастания электротонического потенциала определяется в основном емкостью мембраны. Однако большинство нервных клеток имеют вытянутую форму. Нервное волокно иногда достигает длины 1 м при диаметре 1 мкм. Следовательно, выходя из такой клетки, пропускаемый через нее ток будет распределяться очень неравномерно. Установлено, что по мере увеличения расстояния от источника возбуждения (тока) временной ход электротонического потенциала (электротона) постепенно замедляется. Происходит это потому, что электротон преодолевает сопротивление не только мембраны, но продольное сопротивление внутренней среды самой нервной клетки. Для малых сдвигов потенциала электротонические потенциалы в нерве можно зарегистрировать на расстоянии не более нескольких сантиметров от места их возникновения, т.е. локально.
|
|
Деполяризующий электротонический потенциал, который превышает пороговый уровень, вызывает возбуждение. Возбуждение возможно тогда, когда импульс тока имеет адекватную длительность и амплитуду. Соответственно определенный уровень длительности и амплитуды импульса тока существенно влияет на передачу информации в форме потенциала действия. В этой связи локальный характер деполяризации дендиритов, тел нервных клеток и аксонов различается.
Деполяризация дендритов и соответственно тел нервных клеток наблюдается едва достигается пороговый уровень. Происходит это потому, что деполяризация идет за счет повышения натриевой (+Nа) проницаемости мембраны, которая в дальнейшем продолжает деполяризацию автоматически.

Локальный ответ
Локальный потенциал (ЛП) - это местное нераспространяющееся подпороговое возбуждение, существующее в пределах от потенциала покоя (-70 мВ в среднем) до критического уровня деполяризации (-50 мВ в среднем). Его длительность может быть от нескольких миллисекунд до десятков минут.
1. Возникает в ответ на действие pаздpажителя силой от 0,5 до 0,9 поpога
2. Активная фоpма деполяpизации, поскольку ионная пpоницаемость повышается в зависимости от силы подпоpогового pаздpажителя
3. Гpадуален по амплитуде (амплитуда находится в пpямой зависимости от силы и частоты pаздpажений)
4. Развитие деполяpизации пpоисходит до кpитического уpовня, пpичем не пpямолинейно, а по S-обpазной кpивой. Пpи этом деполяpизация пpодолжает наpастать после пpекpащения pаздpажения, а затем сpавнительно медленно исчезает
5. Способен к суммации (пpостpанственной и вpеменной)
6. Локализуется в пункте действия pаздpажителя и пpактически не способен к pаспpостpанению, т.к. хаpактеpизуется большой степенью затухания
7. Повышает возбудимость стpуктуpы
Виды Локальных ответов(потенциалов):
1. Рецепторный. Возникает на рецепторных клетках (сенсорных рецепторах) или рецепторных окончаниях нейронов под действием стимула (раздражителя). Механизм возникновения такого рецепторного локального потенциала детально рассмотрен на примере восприятия звука слуховыми рецепторами - Молекулярные механизмы рецепции (трансдукции) звука по пунктам Этот процесс называется "трансдукция", то есть преобразование раздражения в нервное возбуждение. Сенсорные рецепторы вторичного типа не умеют порождять нервный импульс, поэтому их возбуждение остаётся локальным и от его амплитуды зависит то, сколько рецепторная клетка выбросит медиатора.
|
|
2. Генераторный. Возникает на сенсорных афферентных нейронах (на их дендритных окончаниях, перехватах Ранвье и/или аксонных холмиках) под действием медиаторов, которые выделили сенсорные клеточные рецепторы вторичного типа. Генераторный потенциал превращается в потенциал действия и нервный импульс при достижении им критического уровня деполяризации, т.е. он генерирует (порождает) нервный импульс. Потому он и назван генераторным.
3. Возбуждающий постсинаптический потенциал (ВПСП). Возникает на постсинаптической мембране синапса, т.е. он отражает передачу возбуждения от одного нейрона к другому. Обычно он составляет +4 мВ. Важно отметить, что возбуждение передаётся от одного нейрона другому именно в виде ВПСП, а не готового нервного импульса. ВПСП вызывает деполяризацию мембраны, но подпороговую, не достигающую КУД и не способную породить нервный импульс. Поэтому обычно требуется целая серия ВПСП для того, чтобы родился нервный импульс, т.к. величина единичного ВПСП совершенно недостаточна для того, чтобы достичь критического уровня деполяризации. Вы можете сами подсчитать, сколько требуется одновременно действующих ВПСП, чтобы родился нервный импульс. (Ответ: 5-6.)
4. Тормозный постсинаптический потенциал (ТПСП). Возникает на постсинаптической мембране синапса, но только не возбуждает её, а, наоборот, тормозит. Соотвтетственно, эта постсинаптическая мембрана входит в состав тормозного синапса, а не возбуждающего. ТПСП вызывает гиперполяризацию мембраны, т.е. сдвигает потенциал покоя вниз, подальше от нуля. Обычно он составляет -0,2 мВ. Используются два механизма создания ТПСП: 1) "хлорный" - происходит открытие ионных каналов для хлора (Cl-), через них в клетку входят ионы хлора и увеличивают её электроотрицательность, 2) "калиевый" - происходит открытие ионных каналов для калия (К+), через них выходят ионы калия, уносят из клетки положительные заряды, что увеличивает электроотрицательность в клетке.
5. Пейсмекерные потенциалы - это эндогенные близкие к синусоидальным периодические колебания мембранного потенциала с частотой 0,1-10 Гц и амплитудой 5-10 мВ. Их генерируют у себя специальные нейроны-пейсмекеры (водителями ритма) самостоятельно, без внешнего воздействия. Пейсмекерные локальные потенциалы обеспечивают периодическое достижение нейроном-пейсмекером критического уровня деполяризации и спонтанную (т.е. самопроизвольную) генерацию им потенциалов действия и, соответственно, нервных импульсов.
Механизм возникновения
Важно понять то, что процесс рождения локального потенциала начинается с открытия ионных каналов. Открытие ионных каналов - это самое главное! Их нужно открыть для того, чтобы в клетку пошёл поток ионов и принёс в неё электрические заряды. Эти ионные электрические заряды как раз и вызывают смещение электрического потенциала мембраны вверх или вниз, т.е. локальный потенциал.
Если открываются ионные каналы для натрия (Na+), то в клетку вместе с ионами натрия попадают положительные заряды, и её потенциал смещается вверх в сторону нуля. Это - деполяризация, и так рождается возбуждающий локальный потенциал. Можно сказать, что возбуждающие локальные потенциалы порождаются натриевыми ионными каналами, когда они открываются.
Образно можно сказать и так: "Каналы открываются - потенциал рождается".
Если открываются ионные каналы для хлора (Cl-), то в клетку вместе с ионами хлора попадают отрицательные заряды, и её потенциал смещается вниз ниже потенциала покоя. Это гиперполяризация, и таким способом рождается тормозный локальный потенциал. Можно сказать, что тормозные локальные потенциалы порождаются хлорными ионными каналами.
Существует также ещё один механизм формирования тормозных локальных потенциалов - за счёт открытия дополнительных ионных каналов для калия (К+). В этом случае из клетки через них начинают выходить "лишние" порции ионов калия, они выносят положительные заряды и увеличивают электроотрицательность клетки, т.е. вызывают её гиперполяризацию. Таким образом, можно сказать, что тормозные локальные потенциалы порождаются дополнительными калиевыми ионными каналами.
Как видите, всё очень просто, главное - открыть нужные ионные каналы. Стимул-управляемые ионные каналы открываются раздражителем (стимулом). Хемо-управляемые ионные каналы открываются медиатором (возбуждающим или тормозным). Точнее, в зависимости от того на какие каналы (натриевые, калиевые или хлорные) будет действовать медиатор, таков будет и локальный потенциал - возбуждающий или тормозный. А медиатор как для возбуждающих локальных потенциалов, так и для тормозных, может быть одним и тем же, тут важно, какие ионные каналы будут связываться с ним своими молекулярными рецепторами - натриевые, калиевые или хлорные.
Потенциал действия
Потенциал действия – это резкое скачкообразное изменение мембранного потенциала с отрицательного на положительный и обратно.
1. Возникает пpи действие pаздpажителей поpоговой и свеpхпоpоговой силы (может возникать пpи суммации подпоpоговых pаздpажителей вследствии достижения уpовня кpитической деполяpизации)
2. Активная деполяpизация пpотекает пpактически мгновенно и pазвивается пофазно (деполяpизация, pеполяpизация)
3. Hе имеет гpадуальной зависимости от силы pаздpажителя и подчиняется закону "все или ничего". Амплитуда зависит только от свойств возбудимой ткани
4. Hе способен к суммации
5. Снижает возбудимость ткани
6. Распpостpаняется от места возникновения по всей мембpане возбудимой клетки без изменения амплитуды
Механизм возникновения
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации. В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется.
Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+ / K+ помпы.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение.
Пороговый потенциал – разность между мембранным потенциалом покоя и критическим уровнем деполяризации. Величина порогового потенциала определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
6. Возбудимость. Изменение возбудимости в процессе возбуждения.
А. Возбудимость клетки во время ее возбуждения быстро и сильно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и, так же как и фазы ПД, определяется состоянием проницаемости клеточной мембраны для ионов. Схематично эти изменения представлены на рис. 3.6.б.
1. Кратковременное повышение возбудимости в начале развития ПД, когда уже возникла частичная деполяризация клеточной мембраны. Если деполяризация не достигает критической величины, то регистрируется локальный потенциал. В случае, если деполяризация достигает Екр, то развивается ПД. При замедленном развитии начальной деполяризации она оценивается как препотенциал. Возбудимость повышена потому, что клетка частично деполяризована, мембранный потенциал приближается к критическому уровню, поскольку открывается часть потенциалчувствительных быстрых Na-каналов. При этом достаточно небольшого увеличения силы раздражителя, чтобы деполяризация достигла Екр, при которой возникает ПД.
2. Абсолютная рефракторная фаза - это полная невозбудимость клетки (возбудимость равна нулю), она соответствует пику ПД и продолжается 1-2 мс; если ПД более продолжителен, то более продолжительна и абсолютная рефракторная фаза. Клетка в этот период при любой силе раздражения не отвечает. Невозбудимость клетки в фазу деполяризации и инверсии (в первую ее половину - восходящая часть пика ПД) объясняется тем, что потенциалзависимые т -ворота Na-каналов уже открыты и ионы Na+ быстро поступают в клетку по всем каналам. Те ворота Na-каналов, которые еще не успели открыться, открываются под влиянием деполяризации - уменьшения мембранного потенциала. Поэтому дополнительное раздражение клетки относительно движения ионов Na+ в клетку ничего изменить не может.


Рис. 3.6. Фазовые изменения возбудимости клетки (б) во время ПД (а). 1,4 - возбудимость повышена; 2 - абсолютная рефрактерная фаза;
2. Относительная рефрактерная фаза - это период восстановления возбудимости, когда сильное раздражение может вызвать новое возбуждение (см. рис. 3.6,5, кривая 3). Относительная рефрактерная фаза соответствует конечной части фазы реполяризации от уровня Екр ± 10 мВ и следовой гиперполяризации клеточной мембраны, что является следствием все еще повышенной проницаемости для ионов К+ и избыточного выхода ионов К+-каналов из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить более сильное раздражение, так как часть Nа+-каналов в конце реполяризации находится еще в состоянии инактивации, а выход ионов К+ из клетки препятствует ее деполяризации. Кроме того, в период следовой гиперполяризации мембранный потенциал больше и, естественно, дальше отстоит от критического уровня деполяризации. Если реполяризация в конце пика ПД замедляется (см. рис. 3.6,а), то относительная рефрактерная фаза включает и период замедления реполяризации, и период гиперполяризации. Рис. 3.6.Фазовые изменениявозбудимости клетки (b) во времяПД (а).1,4-возбудимость повышена;2-абсолютная рефрактерная фаза;3-относительная рефрактерная фаза
4. Фаза экзальтации - это период повышенной возбудимости. Он соответствует следовой деполяризации. В нейронах ЦНС вслед за гиперполяризацией возможна частичная деполяризация клеточной мембраны. В эту фазу очередной ПД можно вызвать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного и оказывается ближе к критическому уровню деполяризации, что объясняют повышенной проницаемостью клеточной мембраны для ионов Nа+. Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность.
Б. Лабильность, или функциональная подвижность (Н.Е.Введенский)— это скорость протекания одного цикла возбуждения, т.е. ПД. Как видно из определения, лабильность ткани зависит от длительности ПД. Это означает, что лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и из клетки, которая, в свою очередь, зависит от скорости изменения проницаемости клеточной мембраны. Особое значение при этом имеет длительность рефрактерной фазы: чем больше рефрактерная фаза, тем ниже лабильность ткани.
Закон силы
Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным - пороговым или сверхпороговым
Учитывая, что порог раздражения является мерой возбудимости, которая определяется порогом деполяризации (разница между потенциалом покоя и уровнем критической деполяризации), то этот закон также должен рассматривать зависимость амплитуды ответа возбудимой ткани от силы раздражителя (раздражитель по силе ниже, равный или выше пороговой величины).
Для одиночных образований (нейрон, аксон, нервное волокно) эта зависимость носит название
Закон "все или ничего"

Подпороговые раздражители не вызывают возбуждение ("ничего"). При пороговых и сверхпороговых воздействиях возникает максимальная ответная реакция ("все"), т. е. возбуждение возникает с максимальной амплитудой ПД
По этому закону также сокращаются сердечная мышца и одиночное мышечное волокно.
Закон не являются абсолютным, а носит относительный характер:
. При действии раздражителей подпороговой силы видимая реакция отсутствует, но возникает местная реакция (локальный ответ)
. При действии пороговых раздражителей растянутая мышечная ткань дает большую амплитуду сокращения, чем не растянутая
При регистрации суммарной активности целостного образования (скелетная мышца, состоящая из отдельных мышечных волокон, нервный ствол, состоящий из множества нервных волокон) проявляется другая зависимость
Чем больше сила раздражителя, тем больше величина ответной реакции
Например
При увеличении силы раздражителя от минимальных (пороговых) до субмаксимальных и максимальных значений амплитуда мышечного сокращения возрастает до определенной величины.
Дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения.
Это связано с тем, что скелетная мышца состоит из множества мышечных волокон, каждое из которых имеет свою возбудимость, а, следовательно, и свой порог раздражения. Поэтому на пороговый раздражитель отвечают только те волокна, которые имеют максимальную возбудимость.
С увеличением силы раздражителя в реакцию вовлекаются все большее количество мышечных волокон, и амплитуда сокращения мышцы все больше увеличивается.
Когда в реакцию вовлекаются все мышечные волокна, составляющие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения
8. Законы раздражения. Закон «силы времени». Понятие о реобазе и хронаксии. Хронаксиметрия и ее клиническое значение.
Закон "силы-времени"

Закон отражает зависимость пороговой силы раздражителя от времени его действия для возникновения возбуждения и гласит:
Возникновение распространяющегося возбуждения зависит не только от силы раздражителя, но и от времени, в течение которого он действует. Чем больше по силе раздражитель, тем меньшее время он должен действовать для возникновения возбуждения
Зависимость носит обратный характер и имеет вид гиперболы. Из этого следует, что на кривой "силы-времени" имеются области, которые не подчиняются этому закону.
Если сила раздражителя будет меньше некоторой (пороговой) величины, то возбуждение не возникнет даже при длительном его воздействии.
Наоборот, если время воздействия будет очень коротким, то возбуждение тоже не возникнет даже при воздействии очень большого по силе раздражителя (в физиотерапии токи высокой частоты используются для получения калорического эффекта)
Для выявления этой зависимости и оценки возбудимости ткани используются следующие количественные характеристики:
Реобаза - это минимальная сила электрического тока, вызывающая генерацию потенциала действия
Полезное время - это минимальное время, в течение которого на ткань должен действовать раздражитель, равный по силе реобазе, чтобы возникло распространяющееся возбуждение
Хронаксия - это минимальное время, в течение которого на ткань должен действовать раздражитель, равный по силе 2 реобазам, чтобы возникло распространяющееся возбуждение
(при поражении нерва хронаксия увеличивается)
ХРОНАКСИМЕТРИЯ (греч. chronos время + axia количество + metreo измерять) - метод определения возбудимости тканей или органов на основе выявления зависимости между пороговой силой электрического раздражения, вызывающего процесс возбуждения, и длительностью его действия. Соответствующие приборы - хронаксиметры состоят из источника постоянного тока, набора сопротивлений и приспособлений для дозирования времени действия электрического тока, подающегося на объект.
Хронаксиметрию применяют в диагностике поражений центральной и периферической нервной системы, опорно-двигательного аппарата, при определении границ патол. очага и функционального состояния отдельных структур головного мозга в процессе проведения нейрохирургических операций. Относительная простота метода и достаточно четкая интерпретация результатов дали возможность использовать хронаксиметрию в спортивной медицине, физиологии труда, промышленной и санитарной гигиене.
Закон градиента

Закон отражает зависимость возникновения возбуждения от скорости или крутизны нарастания силы раздражителя и гласит:
Пороговая сила тока увеличивается при уменьшении крутизны его нарастания до определенной величины. При некоторой минимальной крутизне ответы на раздражение исчезают
Большая крутизна у импульсных токов прямоугольной формы.
Малая крутизна у пилообразных токов (с разным наклоном пилы)
Если сила раздражителя нарастает медленно (длительное действие подпорогового раздражителя), то формируются процессы, препятствующие возникновению ПД.
При этом происходит инактивация Na-каналов.
В результате, нарастание уровня критической деполяризации опережает развитие местных деполяризующих процессов в мембране.
Возбудимость снижается, и порог раздражения увеличивается.
Развивается аккомодация.

Аккомодация - это приспособление ткани к воздействию медленно нарастающего по силе раздражителя, проявляющееся снижением возбудимости
Мер а аккомодации - минимальный градиент или критический наклон
Минимальный градиент - это наименьшая крутизна нарастания тока, при которой раздражающий стимул сохраняет способность генерировать потенциалы действия
Этот показатель также используют для характеристики возбудимости.
Двигательные нервные волокна имеют большую возбудимость, чем скелетная мускулатура.
Поэтому способность к аккомодации, а, следовательно, и минимальный градиент у нервных волокон выше.
Более низкой аккомодацией обладают сенсорные нервные волокна, сердечная мышца, гладкие мышцы, а также образования, обладающие автоматической активностью
Закон аккомодации лежит в основе применения лекарственных препаратов и назначения закаливающих процедур
9. Законы раздражения. Полярный закон. Физиологический электротон. Като-дическая депрессия.
Полярный закон раздражения (закон Пфлюгера)

При замыкании цепи постоянного электрического тока возбуждение возникает только под катодом, а при размыкании - только под анодом
Доказать правильность закона можно при помощи опыта, в котором меняется расположение катода и анода на поврежденном и неповрежденном участках нерва, иннервирующего мышцу
Прохождение постоянного электрического тока через мембрану вызывает изменение мембранного потенциала покоя.
Так, при замыкании цепи возле катода скапливается его "-" заряд, который уменьшает "+" заряд наружной поверхности мембраны.
Разность потенциалов (между наружной и внутренней поверхностями мембраны) уменьшается, и мембранный потенциал изменяется в сторону уровня критической деполяризации, т.е. формируется деполяризация (возбудимость повышается).
Достижение критического уровня приводит к возникновению пикового потенциала (потенциала действия)
При замыкании цепи возле анода скапливается его "+" заряд.
Он увеличивает "+" заряд мембраны и величину мембранного потенциала
Мембранный потенциал удаляется от критического уровня, превышает значение потенциала покоя и формирует гиперполяризацию (возбудимость уменьшается)
При размыкании цепи, прекращение поступления дополнительного "+" заряда от анода приводит к уменьшению (восстановлению) заряда наружной поверхности мембраны.
Мембранный потенциал, уменьшаясь, приближается к критическому уровню
Формируется деполяризация (возбудимость увеличивается).
После достижения критического значения развивается пиковый потенциал
(Поскольку размыкание происходит после замыкания, а, следовательно, на фоне гиперполяризации и пониженной возбудимости, то для возникновения ПД необходим раздражитель, превышающий по силе пороговый - это анодно-размыкательный эффект)
При размыкании возле катода прекращается накопление его "-" заряда
Заряд наружной поверхности мембраны увеличивается (восстанавливается), мембранный потенциал, увеличиваясь, удаляется от критического уровня и возбуждение не возникает
Молекула актина
Это глобулярный белок, состоящий из одного полипептида, который полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга (рис. 4-4 А). Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся мышечном волокне взаимодействие между актином и миозином предотвращают два белка - тропонин и тропомиозин (рис. 4-4 Б).
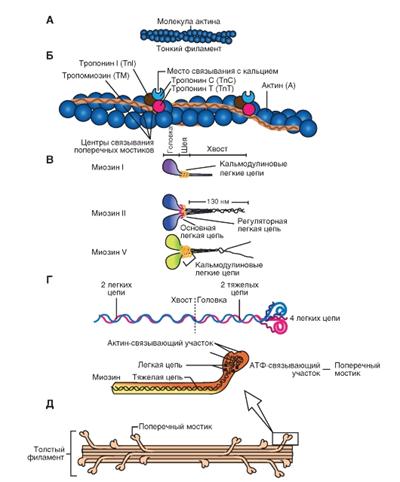
Тропомиозин - стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин - гетеротримерный белок. Он состоит из тропонина Т (отвечает за связывание с одиночной молекулой тропомиозина), тропонина С (связывает ион Са2+) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
Миозин
Это единое название большой семьи протеинов, имеющих определенные отличия в клетках разных тканей. Миозин присутствует у всех эукариотов. Около 60 лет назад было известно два типа миозина, которые сейчас называют миозин I и миозин II. Миозин II был первым из числа открытых миозинов, и именно он принимает участие в мышечном сокращении. Позднее были открыт
|
|
|

Типы оградительных сооружений в морском порту: По расположению оградительных сооружений в плане различают волноломы, обе оконечности...

Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций...

Биохимия спиртового брожения: Основу технологии получения пива составляет спиртовое брожение, - при котором сахар превращается...

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!