

Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...

Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Семя – орган полового размножения и расселения растений: наружи у семян имеется плотный покров – кожура...
Кормораздатчик мобильный электрифицированный: схема и процесс работы устройства...
Топ:
Основы обеспечения единства измерений: Обеспечение единства измерений - деятельность метрологических служб, направленная на достижение...
Когда производится ограждение поезда, остановившегося на перегоне: Во всех случаях немедленно должно быть ограждено место препятствия для движения поездов на смежном пути двухпутного...
Комплексной системы оценки состояния охраны труда на производственном объекте (КСОТ-П): Цели и задачи Комплексной системы оценки состояния охраны труда и определению факторов рисков по охране труда...
Интересное:
Распространение рака на другие отдаленные от желудка органы: Характерных симптомов рака желудка не существует. Выраженные симптомы появляются, когда опухоль...
Национальное богатство страны и его составляющие: для оценки элементов национального богатства используются...
Лечение прогрессирующих форм рака: Одним из наиболее важных достижений экспериментальной химиотерапии опухолей, начатой в 60-х и реализованной в 70-х годах, является...
Дисциплины:
|
из
5.00
|
Заказать работу |
|
|
|
|
История развития биофизики.
Биофизика - молодая наука, сформировавшейся в сер. XX века.
I этап - 17-18в. - накопление отдельных фактов; первые попытки измерений характеристик и параметров био. объектов и систем. Для объяснения использовались законы физики, но делались ссылки на «жизненную силу». Первые идеи биофизики заключались в обосновании материального единства живых организмов и неорг. природы, первых попытках применить физ. законы для объяснения ряда физиологических функции (кровообращения); создан ряд физ. приборов (лупа, микроскоп).
II этап - 18-20в. - проведение экспериментов и опред. многих физ-хим параметров живых организмов. Для объяснения сложных био явлений привлекались законы физики и химии.
III этап - с 20в. - формирование собственного понятийного аппарата, разработка сложных биофиз. методов исследования, выделение ряда разделов в самостоятельные научные дисциплины. Для объяснения био. процессов и явл. привлекают законы математики и биологии.
Стационарное состояние биологических систем.
Состояние системы, при котором ее параметры не изменяются в течение длительного времени, но происходит обмен веществом и энергией с окружающей средой.
Стационарное состояние в живых организмах достигается путем взаимной компенсации всех процессов, связанных с поступлением, удалением и превращением веществ и энергии.
Св-ва стац. состояния системы: стремление системы к мин. ежесекундному приросту энтропии; определеннная внутр. стабильность и упорядоченность.
Принцип Ле-Шателье: если система устойчива, то при отклонении в системе возникают силы, стремящиеся вернуть ее в исходное положение.
Стационарное состояние и термодинамическое равновесие.
|
|
Стац. сосст - сост. системы при кот. ее параметры не изм. в течении длит. времени но происходит обмен веществом и энергией со средой.
ТД равновесие - сост. системы при кот. ее параметры не изм. и она не обменивается с окр. средой ни веществом ни энергией.
Различие между системами:
| Термодинамическое равновесие | Стационарное состояние |
| Отсутствие обмена с окр. средой веществом и энергией | Непрерывный обмен с окр. средой веществом и энергией |
| Энтропия постоянна и соотв. max возможному в данных условиях значению | Энтропия постоянна, но не равна max возможному в данных условиях значению |
| Полное отсутствие в системе каких-либо градиентов | Наличие постоянных по величине градиентов |
| Не требуется затраты свободной энергии | Необходимы постоянные затраты энергии |
| Система нереакционноспособна и не совершает работу против внешних сил | В системе совершаются необратимые реакции, ее работоспособность постоянна и не равна нулю |
Живой организм может изменить уровень стационарного состояния в результате воздействия окр. среды и при патологических процессах.
Одной из важнейших характеристик био. систем является устойчивость стационарных состояний, при отклонении системы от стационарного уровня в ней возникают силы, стремящиеся вернуть ее в первоначальное положение.
В момент смерти организм находится в сост. ТД равновесия
Уравнение Пригожина.
в стац. состоянии при фиксированных внеш. параметрах скорость продукции энтропии в открытой системе, обусловленная протеканием необратимых процессов, постоянна во времени и минимальна по величине:
diS/dt → min
Т.о. стац. состояние характеризуется мин. рассеянием энергии. В соответствии с теоремой Пригожина для поддержания стац. состояния тратится некоторое кол-во энергии, причем самое минимальное. Организм стремится работать на самом выгодном энергетическом режиме. При этом энтропия возрастает с минимальной скоростью.
|
|
Основные понятия и особенности кинетики биологических процессов.
базируются на общих теоретических положениях хим. кинетики, изучающей скорости хим. реакций. Фундаментальное понятие хим. кинетики - хим. реакция, представляющая собой совокупность актов перегруппировки межатомных связей.
Хим. реакции можно разделить на:
1) Гомогенные протекают с одинаковой скоростью в любом элементарном объеме данной фазы;
2) гетерогенные - на границе раздела фаз, и скорость их определяется скоростью подачи реагирующих веществ на поверхности раздела фаз.
Катализаторы повышают скорость спонтанно протекающих реакций. Если вещество инициирует реакцию, оно называется инициатором, если ускоряет каталитическую реакцию, то его относят к активаторам. Соединения, понижающие скорость каталитической реакции или полностью подавляющие - ингибиторами.
Под кинетикой реакции понимают зависимость скорости реакции от концентрации реагирующих веществ, температуры и других параметров.
Реакции называются последовательными, если продукт одной из реакций является исходным веществом для другой. A 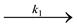 B
B  C.
C.
Сопряженной называют реакцию, которая происходит при одновременном протекании другой реакции.
Цепная реакция - это самоподдерживающаяся химическая реакция, при которой первоначально появляющиеся продукты принимают участие в образовании новых промежуточных и конечных продуктов.
Параллельными называют совместно протекающие реакции, если, по крайней мере, одно исходное вещество этих реакций является общим.
Аллостерические ферменты.
Аллостерический механизм – контроль активности фермента реализуется путем изм. конформации белковой молекулы в результате связывания метаболита–регулятора в особом – аллостерическом центре, пространственно удаленном от активного центра.
Характерной особенностью аллостерических ферментов является наличие нескольких активных и нескольких аллостерических центров в молекуле фермента. Это связано с тем, что аллостерические ферменты, как правило, обладают четвертичной структурой, то есть являются олигомерами, состоящими из нескольких субъединиц.
Молекулы аллостерических ферментов содержат набор активных центров и набор аллостерических или регуляторных центров.
Аллостерические ферменты часто выполняют ключевую роль в регуляции обмена веществ, поскольку обладают способностью определять количество важных метаболитов и изменять в соответствии с этим свою активность.
|
|
Понятие о фолдинге белков
В биохимии и молекулярной биологии фолдингом белка (укладкой белка) называют процесс спонтанного сворачивания полипептидной цепи в уникальную нативную пространственную структуру (так называемая третичная структура).
В совокупности молекулы, участвующие в фолдинге белков, называют регуляторами фолдинга, среди которых выделяют несколько типов. Молекулы, ускоряющие фолдинг, называются катализаторами фолдинга. Молекулы, служащие для изменения формы белка, — шаперонами фолдинга. Существует четыре типа молекул, которые играют роль таких шаперонов.
1. Молекулы, обеспечивающие правильный фолдинг белков (фолдинг-шапероны).
2. Молекулы, созданные для удержания частично свернутой молекулы белка в определенном положении. Это необходимо, чтобы система имела возможность закончить фолдинг (удерживающие шапероны).
3. Шапероны, разворачивающие белки с неправильной формой (дезагрегирующие шапероны).
4. Шапероны, сопровождающие белки, транспортируемые через клеточную мембрану (секреторные шапероны).
Фолдинг шапероны помогают белку принять правильную конформацию. Многие из них являются небольшими сахарами или пептидами. В секреции белков из клетки участвует другая контролирующая система, которая включает секреторные шапероны. Секреторные шапероны узнают сигнальную последовательность аминокислот, которую соответственно называют секреторной последовательностью. Эта последовательность связывается с секреторным шапероном, шаперон поступает внутрь мембраны, обеспечивая экспорт белка вместе с собой.
Денатурация белков
Сравнительно слабые связи, ответственные за стабилизацию вторичной, третичной и четвертичной структур белка, легко разрушаются, что сопровождается потерей его биологической активности. Разрушение исходной (нативной) структуры белка, называемое денатурацией, происходит в присутствии кислот и оснований, при нагревании, изменении ионной силы и других воздействиях. Как правило, денатурированные белки плохо или совсем не растворяются в воде. При непродолжительном действии и быстром устранении денатурирующих факторов возможна ренатурация белка с полным или частичным восстановлением исходной структуры и биологических свойств.
|
|
Как правило, белки достаточно стабильны в тех условиях (температура, pH и др.), в которых они в норме функционируют в организме. Резкое изменение этих условий приводит к денатурации белка. В зависимости от природы денатурирующего агента выделяют механическую (сильное перемешивание или встряхивание), физическую (нагревание, охлаждение, облучение, обработка ультразвуком) и химическую (кислоты и щёлочи, поверхностно-активные вещества, мочевина) денатурацию.
Денатурация белка может быть полной или частичной, обратимой или необратимой. Денатурация в некоторых случаях обратима, как в случае осаждения водорастворимых белков с помощью солей аммония, и используется как способ их очистки.
20.Биофизика белка. Динамические свойства белка.
В пептидной цепи им-ся 2 типа степеней свободы:
1. высокочастотные колебания (1013 с-1) хар-ся смещением атомов на доли ангстрем ~ 0,1А волновых колебаний длин жестких связей валет-х узлов.
2. ню ~ 106–109 с-1 Амплитуда смещения ~ 1-2 А (обусловлены): степени свободы связывание с вращением 1-х связей. Е-го затратный: 2-5 ккал/моль на поворот полного угла именно эти колебания обеспеч-ют конформацию белка.
Растворимость белков
Белки бывают растворимые и нерастворимые в воде. Растворимость белков зависит от их структуры, величины рН, солевого состава раствора, температуры и других факторов и определяется природой тех групп, которые находятся на поверхности белковой молекулы. К нерастворимым белкам относятся кератин (волосы, ногти, перья), коллаген (сухожилия), фиброин (щелк, паутина). Многие другие белки растворимы в воде. Растворимость определяется наличием на их поверхности заряженных и полярных группировок (-СОО-, -NH3+, -OH и др.). Заряженные и полярные группировки белков притягивают к себе молекулы воды, и вокруг них формируется гидратная оболочка существование которой обусловливает их растворимость в воде.
LgCin / lgCout)
Ем –равновесный потенциал, определяемый как разностьпотенциалов по обе стороны мембраны Т – температура (по Кельвину), F – число Фарадея, R- универсальная газовая постоянная
Cin и Cout – концентрация потенциалопределяющих ионов по обе стороны мембраны.Для более точного вычисления величины мембранного потенциала необходимо учитывать диффузию ионов К, Na, Cl. В связи с этим для определения мембранного потенциала используют уравнение Гольдмана:
|
|
lg
Pk,na,cl – коэффициенты проницаемости для соответствующих ионов внутри и снаружи клетки.
В состоянии покоя проницаемость мембраны для ионов калия больше, чем для натрия и больше, чем для хлора: Pk ≥ PNa, Pk > Pcl.
Урвнение Нернста и Гольдмана не учитывают активного транспорта ионов через мембрану, наличие в мембране ионных насосов. В мембране функционируют молекулы Na, K- АТФазы, перекачивающие калий внутрь, натрий из клетки. С учетом работы ионных насосовиспользуют уравнение Томаса:
lg
m – отношение количества ионов натрия к ионам калия, перекачиваемых ионными насосами через мембрану. Чаще всего Атфаза работает в режиме, когда m = 3/2.
Цитоплазма клетки в состоянии покоя всегда имеет отрицательный потенциал по отношению к межклеточной жидкости.
Спектральные св-ва белков.
Гемопротеиды. Состоит из 2х частей – гем + белок – глобин. Сущ. различные типы гемоглобина. У гемма 4 кольца соед-х с Fe2+. В зав-ти от того в каком вален-м состоянии находится Fe гемма мол-ла будет иметь спектр-ые хар-ки.
Лиганд – соед, кот координирует с атомом метала. Кроме О2 в кач лиганда мог выступать др гр-пы. НbСО – карбоксигемоглобин.
Спектр-ые хар-ки НвО2
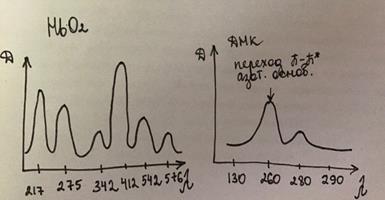
Полосы поглощ-ия до 300нм относят к глобину, после 300нм – к гемму.
В спектре поглощения НвО2 следует обратить внимание на – 275 и 412.
Пол. погл с мах 412 наз пол. Соре.
Пол. погл-ия гем-на с λмах = 275нм обусловл светопоглощ за счет π→π* разрыхляющ.
Речь идет об электронных переходах за счет ненасыщ связи в ароматических соединениях
ДНК – материальный субстрат наслед-ти, квази-кристал-ая молекула, упорядоченная. Спектр погл ДНК форм-ся за счет спектрал-х св-в нуклеотидов, а спектр погл нуклеотидов форм-ся за счет спектра погл азот оснований. В состав ДНК входят пурин-е и(А,Г) и пиримид-ые (Т, Ц,) азот основ-я.
Дозиметрия.
Дозиметрия - изучают величины, характеризующие действие ионизирующего излучения на вещества, а также методы и приборы для их измерения.
Поглощенная доза излучения - это энергия ионизирующего излучения, поглощенная единицей массы поглощающего вещества. Поглощенная доза определяется для всех видов ионизирующего излучения. Она зависит от природы излучения и свойств вещества. Поглощенная доза измеряется в Греях (Гр). 1 Грей – доза, которая характеризует поглощение 1 килограммом вещества 1 Джоуля энергии.
Экспозиционная доза – это общее количество радиоактивного излучения, достигающего вещества. не зависит от характеристик вещества, а определяется только характеристиками излучения.
Энергия ионизирующих излучений значительно отличается от тепловой энергии. Смертельная экспозиционная доза гамма-лучей очень незначительно изменяет температуру тела. Излучения, проходя через живые клетки, изменяют структуру важных био молекул. При этом длительное действие небольшой дозы излучения является более благоприятным, чем быстрое действие той же дозы, поскольку при длительном действии происходят исправления (репарация) некоторых повреждений
Био эффект действия радиации при прочих равных обстоятельствах прежде всего определяется дозой облучения.
Измерение дозы излучения с целью предсказания радиационного эффекта осуществляют с помощью дозиметрических приборов - дозиметров.
В целях регламентации (гигиенического нормирования) радиационного воздействия на человека пользуются понятием предельно допустимой дозы за год (ПДД). Это – наибольшая величина индивидуальной эквивалентной дозы за год, которая при равномерном воздействии в течение 50 лет не вызовет в состоянии здоровья персонала (профессиональных работников) неблагоприятных изменений, обнаруживаемых современными радиобиологическими методами. Предельную эквивалентную дозу за год для ограниченной части населения называют пределом дозы за год (ПД).
История развития биофизики.
Биофизика - молодая наука, сформировавшейся в сер. XX века.
I этап - 17-18в. - накопление отдельных фактов; первые попытки измерений характеристик и параметров био. объектов и систем. Для объяснения использовались законы физики, но делались ссылки на «жизненную силу». Первые идеи биофизики заключались в обосновании материального единства живых организмов и неорг. природы, первых попытках применить физ. законы для объяснения ряда физиологических функции (кровообращения); создан ряд физ. приборов (лупа, микроскоп).
II этап - 18-20в. - проведение экспериментов и опред. многих физ-хим параметров живых организмов. Для объяснения сложных био явлений привлекались законы физики и химии.
III этап - с 20в. - формирование собственного понятийного аппарата, разработка сложных биофиз. методов исследования, выделение ряда разделов в самостоятельные научные дисциплины. Для объяснения био. процессов и явл. привлекают законы математики и биологии.
Современные проблемы биофизики.
1) выявление молекулярной организации биополимеров и ее связи со свойствами и выполняемыми функциями.
2) выявление структуры и функционирования молекулярных и мембранных машин (лекарственных препаратов)
3) изучение молек. датчиков, рецепторов, путей и механизмов утилизации в био.молекулахэнергии фосфорных соединений.
4) исследование фотобиолгической, рабиобиолог., первыичных механизмов при поглащении внешней энергиибио системами.
4. Термодинамика биологических процессов, основные понятия.
это раздел биофизики, изучающий закономерности превращения энергии в биосистемах на макромолекулярном уровне. Законы термодинамики исп. в биологии для расчета параметров энергетических превращений в организме и для определения КПД био. процессов.
Термодинамическая система - часть пространства ограниченная поверхностью раздела (клетка, митохондрия). Существует 3 типа систем, отличающихся по характеру взаимодействия с внешней средой.
1) Изолированные - не обмениваются с окр. средой ни веществом, ни энергией, т.е. границы такой системы непроницаемы.
2) Закрытые - обменивающиеся через свои границы энергией с окр. средой, но непроницаемые для веществ.
3) О ткрытые - осуществляется обмен веществом и энергией.
Термодин. системы характеризуются опр. свойствами или термодин. параметрами:
1) не зависят от массы (температура, давление) - интенсивные.
2) зависящие от количества вещества или массы - экстенсивными (объем, число молей).
Совокупность изм. состояний в системе - термодинамическим процессом, которые делят на:
1) Обратимыми - при которых возвращение системы в первоначальное состояние не требует затрат энергии извне; не происходит рассеивания энергии в виде теплоты.
2) Необратимыми - возвращение системы в исходное состояние возможно лишь при условии затрат внешней энергии; характеризуются переходом части энергии в теплоту. Термодин. био процессов изучает необратимые (неравновесные) процессы.
5. Тепловые эффекты в биосистемах.
1 закон ТД - закон сохр. энергии: в изолированной термодин. системе полный запас энергии есть величина постоянная и возможны только превращения одного вида энергии в др. в эквивалентных соотношениях: U = const; ΔU = 0.
для закрытых систем: теплота, подведенная к системе, расходуется на изм. внутр. энергии системы и на совершение работы против внешних сил: δQ = dU + δA,
(Q - теплота, подведенная к системе; U - внутр. энергия системы; А – работа; δ обозначает, что теплота и работа не явл. функциями состояния системы и не могут быть полными дифференциалами).
Жизнедеятельность организмов сопровождается выделением в окр. среду тепловой энергии за счет дыхания, пищеварения, работы мышц. Теплота, выделенная организмом, может быть разделена на первичную (основную) и вторичную (активную).
Первичная теплота - результат неизбежного рассеивания энергии из-за необратимо протекающих биохимических реакций. Первичная теплота выделяется сразу же после поглощения организмом кислорода и продуктов питания независимо от того, совершает он работу или нет. Она идет на нагревание организма и рассеивается в окружающем пространстве.
Выделение вторичной теплоты наблюдается лишь при реализации энергии макроэргических соединений (АТФ, ГТФ) в ходе обменных реакций или процессов жизнедеятельности.
Для живых организмов в нормальных условиях характерно равновесие между первичной и вторичной теплотой. При патологических процессах увеличивается доля первичной теплоты, что сопровождается повышением температуры тела.
Закон Гесса - тепловой эффект хим. реакции, развивающейся через ряд промежуточных стадий, не зависит от пути перехода, а определяется лишь разностью энтальпий конечных и исходных продуктов реакции.
Q = ΔН = ∑Hр-∑Hс
(где ∑Hр - сумма энтальпий всех продуктов реакции; ∑Hс - сумма энтальпий всех исходных веществ; ΔН - изменение энтальпии химической реакции).
Второе начало “Теплота не может переходить сама собой от более холодного тела к более теплому”.
|
|
|

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни...

История создания датчика движения: Первый прибор для обнаружения движения был изобретен немецким физиком Генрихом Герцем...

Архитектура электронного правительства: Единая архитектура – это методологический подход при создании системы управления государства, который строится...

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰)...
© cyberpedia.su 2017-2024 - Не является автором материалов. Исключительное право сохранено за автором текста.
Если вы не хотите, чтобы данный материал был у нас на сайте, перейдите по ссылке: Нарушение авторских прав. Мы поможем в написании вашей работы!